|
魚類用語
![]() イソギンチャクと共生する色彩豊かなカクレクマノミ(スズメダイ科)。 イソギンチャクと共生する色彩豊かなカクレクマノミ(スズメダイ科)。
本項、魚類用語(ぎょるいようご)では、魚類の身体の名称、特徴や生態などを示す用語について記す。ただし、狭義の魚類に含まれない無顎類(円口類)を含む。
身体の構造
ウィキメディア・コモンズには、 魚類解剖学に関連するカテゴリがあります。
 サメの体の構造。体部は頭部、軀幹部、尾部に分けられる。 サメの体の構造。体部は頭部、軀幹部、尾部に分けられる。
魚類の体部は頭部、軀幹部、尾部に分けられる。
頭部
ウィキメディア・コモンズには、 頭部に関連するカテゴリがあります。
- 頭部(とうぶ、英: head)
- 吻端から鰓蓋末端までの部分。
- 吻(ふん、英: snout)
- 眼の前縁より前の部分。吻の形は食性によって異なる。ヒイラギ、マトウダイなどは膜を折りたたんだような吻を一気に伸ばして獲物を捕らえる。ヤガラ類などは吻部が筒状に伸び、カジキ、サヨリなどは吻部が槍のように延長する。吻の先端は吻端(ふんたん)と呼ばれる。
- 眼隔域(がんかくいき)
- 眼と眼の間。
- 口裂(こうれつ)
- 口の開口部。
- 額部(がくぶ、英: frontal region)
- 眼の背後方に当る頭部。
- 頬部(きょうぶ、英: cheek)
- 眼より下方の部位。眼と前鰓蓋骨隆起線との間。
- 後頭部(こうとうぶ、英: occipital region) または 後頭(こうとう)
- 眼より後方の頭部背面。項部の直前の部分。後頭部は魚種により突出したり、窪んだり、棘や皮弁を備えたりする。
- 項部(こうぶ、英: nape)
- 後頭部直後の部分。軀幹部近くの頭部背面。後頭部などとの境界は厳密には定められていない。
- 縫合部(ほうごうぶ、英: symphysis) または 下顎縫合部(かがくほうごうぶ)
- 両側の下顎が合着する部位。
- 頤(おとがい、英: chin, mentum) または 頤部(おとがいぶ、英: mental region、 羅: regio mentalis)
- 縫合部の後方部。下顎縫合部の直後。
- 峡部(きょうぶ、英: isthmus)
- 左右の鰓膜が接近する付近の頭部下面。頤の後方。
- 喉部(こうぶ、英: jugular)
- 峡部の直後、胸部との間。胸鰭基底の下部より前の部分。
軀幹部
- 軀幹部(躯幹部、くかんぶ、英: trunk) または 胴(どう)、胴部(どうぶ)
- 鰓蓋(または鰓孔)末端から肛門(または総排泄孔)までの部分。大きく胸部と腹部に分けられるが、これらの境目に明確な区別はない。
- 肩部(けんぶ、英: shoulder)
- 軀幹部の前側部、胸鰭を支える骨(肩帯)があるところ。
- 胸部(きょうぶ)
- 軀幹部の下面前方。鰓蓋末端から腹部前端までの軀幹部。喉部の後方。
- 腹部(ふくぶ、英: abdomen)
- 軀幹部の下面後方。胸部後端から肛門までの軀幹部。
- 腋(えき、英: axil) または 腋部(えきぶ)
- 対鰭(普通胸鰭)内部の基底付近の体部。腹鰭の腋部として使用されることもある。
尾部
ウィキメディア・コモンズには、 尾部に関連するカテゴリがあります。
- 尾部(びぶ、英: tail)
- 肛門より後方の体部。肛門から尾鰭基底まで。
- 尾柄(びへい、英: caudal peduncle) または 尾柄部(びへいぶ)
- 尾部のうち臀鰭基底末端(臀鰭の最後の軟条基底)から尾鰭起部までの部分。
鰭
ウィキメディア・コモンズには、 鰭に関連するカテゴリがあります。
 一般的な魚類(カクレクマノミ)の鰭。 一般的な魚類(カクレクマノミ)の鰭。
- 鰭(ひれ、英: fin、羅: pinna, pterygium)
- 魚の体壁から突出する扁平な構造。推進力を産み出し、体の安定を担う運動器官。鰭条、鰭膜からなる。また、対鰭と不対鰭に大別される。
- 鰭条(きじょう、英: fin ray) または 条(じょう)
- 鰭の支持物として担鰭骨から鰭の外縁に向かって放射状もしくは平行に走る細線状の構造。鰭膜とともに鰭を形成する。ホウボウの胸の付け根にある「足」は味覚を感じる器官だが、これは鰭条の一部が変化したものである。
- 角質鰭条(かくしつきじょう、英: ceratotricha)
- 軟骨魚類の鰭条。角質からなり、分節せず細長い。
- 鱗状鰭条(りんじょうきじょう、英: lepidotrichia)
- 硬骨魚類の鰭条。骨質からなり、細かく分節する。軟条と棘条に分かれる。
- 棘条(きょくじょう、英: spinous ray) または 棘(きょく、英: spine)、鰭棘(ききょく)
- 固く、分節していない鱗状鰭条。軟条より骨化が進んでいる。左右の接合分節が認められない。全ての鰭に棘条がない硬骨魚類もいる(原棘鰭上目など)。背鰭のものを背鰭棘(せびれきょく)、臀鰭のものをものを臀鰭棘(しりびれきょく)という。アンコウやカエルアンコウのように擬餌状体と誘引突起のように変化したものや、ゴンズイやオコゼ、アイゴなどのように毒腺を持つものもある。
- 遊離棘(ゆうりきょく)
- 一つの鰭の中で他の鰭条から離れている棘。
- 棘状軟条(きょくじょうなんじょう、英: spine soft ray)
- 普通棘条は分節しないが、コイ目の背鰭、臀鰭前部には分節的な棘条がある。
 大きな胸鰭と指状の軟条をもつホウボウ(ホウボウ科)。 大きな胸鰭と指状の軟条をもつホウボウ(ホウボウ科)。
- 軟条(なんじょう、英: soft ray)
- 柔軟で分節している鱗状鰭条。分節のある左右1対の鰭条が癒合した状態。多くの場合先端は癒合せずに分節している。
- 分枝軟条(ぶんしなんじょう、英: branched soft ray)
- 起部が1本で先端に向かって分枝する軟条。分枝条(ぶんしじょう)、分岐軟条(ぶんきなんじょう)とも呼ばれる。
- 不分枝軟条(ふぶんしなんじょう、英: unbranched soft ray)
- 起部から先端まで1本の軟条。不分枝条(ふぶんしじょう)、不分岐軟条(ふぶんきなんじょう)とも呼ばれる。
- 遊離軟条(ゆうりなんじょう、英: free soft ray)
- 一つの鰭の中で他の鰭条から離れている軟条。多くはほかのものより太く、感覚能がある。
- 鰭膜(きまく、英: fin membrane)
- 鰭条を連結し、鰭を形成する膜。 鰭条間膜(きじょうかんまく)、条間膜(じょうかんまく)とも呼ばれる。
- 基底(きてい、英: fin base, base of fin)
- 鰭の前端基部から後端基部までの部分。
- 起部(きぶ、英: origin of fin) または 始部(しぶ)
- 鰭の最前端起部。
- 対鰭(ついき、ついびれ、英: paired fin) または 有対鰭(ゆうついき)
- 魚類の両体側にあって対となっている鰭。胸鰭と腹鰭を言う。水平翼として揚力を生じさせると同時に体のローリングを防ぐ。遊泳にブレーキをかけ、進行方向に対して左右、上下に方向転換する際のバランスを取る。
 ナンヨウマンタ(トビエイ科)の頭鰭。 ナンヨウマンタ(トビエイ科)の頭鰭。
- 頭鰭(とうき、あたまびれ、英: cephalic fin)
 胸鰭と腹鰭を拡げて飛翔するトビウオ(トビウオ科)。 胸鰭と腹鰭を拡げて飛翔するトビウオ(トビウオ科)。
- 胸鰭(むなびれ、きょうき、英: pectoral fin)
- 胸に対にある鰭。四肢動物の前肢に当る。トビウオでは飛行に使用される。ホウボウ科では胸鰭が著しく長く、下部の軟条3対は肥厚して遊離し、指状の動きをして餌生物を探す。ウツボ(ウナギ目)やタウナギ(タウナギ目)にはない。
- 骨質盤(こつしつばん、英: Lamina Circularis)
- ドジョウ科特有のもので、雄の胸鰭基部に発達する。ドジョウ科の種の判別は骨質盤の形で判別する。
- 腹鰭(はらびれ、ふっき、英: pelvic fin, abdominal fin)
- 腹に対にある鰭。四肢動物の後肢に当る。サメの腹鰭は交尾器(鰭脚)、ハゼの腹鰭は吸盤に変化する。またウツボやウナギ(ともにウナギ目)、タウナギ(タウナギ目)にはない。
- 腹鰭前葉(はらびれぜんよう、ふっきぜんよう)
- ガンギエイなどが持つ。腹鰭の前部。
- 不対鰭(ふついき、英: unpaired fin) または 正中鰭(せいちゅうき、英: median fin)、垂直鰭(すいちょくき、すいちょくびれ)、主鰭(しゅき)、無対鰭(むついき)
- 体の正中線上にある鰭。背鰭、臀鰭、尾鰭、小離鰭の総称。体の横揺れを防ぎ、泳ぎの推進力を生み出し、方向転換を補助する。
 多基の背鰭を持つポリプテルス・デルヘッツィ(ポリプテルス科)。 多基の背鰭を持つポリプテルス・デルヘッツィ(ポリプテルス科)。
- 背鰭(せびれ、はいき、英: dorsal fin、羅: pinna dorsails)
- 背中線上に沿って1–3基ある鰭。遊泳の際、体の安定性を保つ。また瞬間的な方向転換に役立つ。1基が普通であり、マサバ、カツオ、ブリ、マアジ、カワハギなどは2基、ヘビギンポ、マダラ、スケトウダラなどは3基、ポリプテルスなどは数基ある。
- 第1背鰭(第一背鰭、だいいちせびれ、英: first dorsal fin) または 棘条背鰭(きょくじょうせびれ)
- 背鰭の第1基。ほとんどの魚類が持つ。
- 第2背鰭(第二背鰭、だいにせびれ、英: second dorsal fin) または 軟条背鰭(なんじょうせびれ)
- マサバ、カツオ、ブリ、マアジ、カワハギなどがもつ。
- 第3背鰭(第三背鰭、だいさんせびれ、英: third dorsal fin)
- タラ類などが有する。
- 臀鰭(しりびれ、でんき、尻鰭、英: anal fin)
- 遊泳の際、体の安定性を保つ。遊泳そのものに重要な役割はない。穴を掘って産卵床を作る魚類においては、雌が産卵床を掘る際、礫と礫の間に砂利や砂を除去できたかどうかを探る道具として使われる。通常1基だが、リュウグウノツカイ、フリソデウオなどは0基、タラ類は2基ある。
- 臀鰭様膜突起(しりびれようまくとっき)
- ヤツメウナギが持つ鰭状突起物
 メバチ(サバ科)の尾部。finlets=小離鰭とkeels=尾柄隆起(キール)。 メバチ(サバ科)の尾部。finlets=小離鰭とkeels=尾柄隆起(キール)。
- 尾鰭(おびれ、びき、英: caudal fin、羅: pinna caudails)
- 魚類の尾端にあり、脊柱後端部で支持される鰭。支持する骨格との関係で分類される。左右の動き(ビート)で遊泳時の主な推進力を作り出す。尾鰭の上部を上葉(じょうよう)、下部を下葉(かよう)という。リボンイワシの幼魚期の尾鰭は著しく長く、尾鰭長(リボン状尾鰭)は体長の14.6倍。
- 原正尾(げんせいび、英: diphycercal tail) または 両尾(りょうび)
- 尾鰭後端が細く尖り、脊椎骨末端がまっすぐな尾鰭。背腹で相称となる。円口類などがもつ、原始的な尾鰭である。発生的には下記の尾鰭も原生尾に近い形態を経過して形成される。
- 異尾(いび、英: heterocercal tail) または 歪尾(わいび)
 マオナガ(オナガザメ科)。尾鰭上葉が著しく長い。これを使い餌となる小魚を弾いて弱らせ、捕食する。 マオナガ(オナガザメ科)。尾鰭上葉が著しく長い。これを使い餌となる小魚を弾いて弱らせ、捕食する。
- 上葉と下葉が著しく異なった尾鰭。軟骨魚類、チョウザメなど古いグループのもの。脊柱の尾端が背側に曲がってできるため、サメの尾鰭の上葉の中には脊柱が走り末端まで通っているのに対し、下葉は鰭を支える下索軟骨と角質鰭条で支持されているだけで柔らかい。上葉は縮小し、下葉が発達するため、背腹で不相称となる。
- 正尾(せいび、英: homocercal tail)
- 上下両尾が同長で同形な尾鰭。真骨類の多くが持つ。異尾よりもいっそう上葉が縮小し、下葉が発達して相称となることで形成される。
- 橋尾(きょうび、英: gephyrocercal tail)
- ウナギ、ハイギョにみられるように、二次的に原正尾に近い形になったもの。マンボウの舵鰭もこの名で呼ばれることがある。
- 擬尾(ぎび)
- タラ科魚類にある特殊な尾鰭骨格。脊椎骨の後方の神経棘と血管棘が尾鰭棘条の大部分を支持する。
- 尾鰭後縁(おびれこうえん、びきこうえん)
- 尾鰭の最後の縁。尾鰭後縁の形は種によってさまざまである。生態と関係が深い。円いものを円形(えんけい、英: rounded)、まっすぐなものを截形(せつけい、英: truncate)、円く窪んでいるものを湾入形(わんにゅうけい、わんにゅうがた、英: emarginate)や三日月形(みかづきけい、みかづきがた、英: lunate)、鋭く切れ込んでいるものは二叉形(にさけい、英: forked)、後方に鋭く突出するものは尖形(せんけい)と呼ばれる。楔形(くさびがた、英: wedge-shaped)、二重湾入形(にじゅうわんにゅうけい、にじゅうわんにゅうがた)、二重截形(にじゅうせつけい)のものもある。截形や楔形のものは泳ぎが遅く、三日月形や二叉形のものは泳ぎが速い。マダイは二叉形、ジンベエザメは三日月形の尾鰭を持つ。
- 膜鰭(まくき、英: fin fold)
- 多くの種の不対鰭は、発生過程で一続きの膜状の鰭となることがあり、この膜状の鰭をいう。幼生期に中間部が消失して鰭条を持った成魚の鰭に置き換わる。
 サケ科魚類の鰭。1.鰓蓋、2.側線、3.背鰭(棘条はない)、4.脂鰭、5.尾鰭、6.臀鰭、7.腹鰭、8.胸鰭 サケ科魚類の鰭。1.鰓蓋、2.側線、3.背鰭(棘条はない)、4.脂鰭、5.尾鰭、6.臀鰭、7.腹鰭、8.胸鰭
- 脂鰭(あぶらびれ、英: adipose fin)
- 背鰭と尾鰭の中間にある膜状の小さな鰭状突起物。原則として鰭条が欠く。機能は定かではない。真骨類の低位グループにある。旧サケ目(サケ目のサケ科、キュウリウオ目のキュウリウオ科やシラウオ科)、ハダカイワシ目のハダカイワシ科、ナマズ目のハマギギ科(ゴンズイ科などにはない)、カラシン目、ヒメ目のエソ科、アオメエソ科など約10科がもつ。
- 小離鰭(しょうりき、英: finlets) または 離鰭(りき、はなれびれ)
- 背鰭や臀鰭後方に独立にある、小さな遊離した鰭。泳ぐ際、水の渦流を少なくする。マグロ、アジ、サンマ、サワラなどがもつ。
- イトマキエイ類(イトマキエイ、オニイトマキエイ、ヒメイトマキエイなど)の頭部にある耳状の突起。体盤を形成する胸鰭が頭の両側でいったん消失し、吻端で再度出現し形成される耳朶状のもの。近縁のトビエイ類にはない。食事の際手や唇のような働きをする。
 マンボウ(マンボウ科)の骨格。舵鰭は背鰭と臀鰭の後方の変形物であることがわかる。 マンボウ(マンボウ科)の骨格。舵鰭は背鰭と臀鰭の後方の変形物であることがわかる。
- 舵鰭(かじびれ、英: clavus) または 橋尾(きょうび)
- マンボウの体の後端にある鰭。マンボウには尾鰭がなく、背鰭と臀鰭の要素が組み合わさってできたもの。左右に舵を切る。
鱗
ウィキメディア・コモンズには、 鱗に関連するカテゴリがあります。
 硬鱗(ガノイン鱗)で覆われたコチョウザメ(チョウザメ科) 硬鱗(ガノイン鱗)で覆われたコチョウザメ(チョウザメ科)
 コイ(コイ科)の円鱗。 コイ(コイ科)の円鱗。
 ブリーム(コイ科)の円鱗。 ブリーム(コイ科)の円鱗。
- 鱗(うろこ、英: scale)
- 体表を覆う硬質の小薄片状形成物。真皮に起生する皮骨(外骨格)。大きく楯鱗、硬鱗、骨鱗に区別される。
- 楯鱗(じゅんりん、英: placoid scale) または 皮歯(ひし、英: dermal tooth, dermal denticle)
- 板鰓類の鱗。真皮に形成され、骨性の基底版から突起が体表に突出する。突起内部には髄腔があり、血管や神経が入る。髄腔を象牙質が囲み、外表面は表皮由来のエナメロイド(英: enameloid、中胚葉性エナメル、ちゅうはいようせいエナメル、英: mesodermal enamel)が覆う。歯と同じ構造をしているため皮歯とよばれるが、発生も歯と同様である。サメ類では全身に密生し「鮫肌」となるが、エイ類では退化的で体表各所に点在する。とくにシビレエイは完全に退化する。
- 硬鱗(こうりん、英: ganoid scale)
- 狭義にはガノイン鱗のこと。広義にはコズミン鱗を含む。
- ガノイン鱗(ガノインりん、英: ganoid scale, rhomboid scale) または 硬鱗(こうりん)
- ガー、チョウザメ類などがもつ鱗。真皮内の骨板中に細い血管が走り、その表面は表皮と真皮に由来するガノイン(英: ganoin、硬鱗質、こうりんしつ)で覆われる。ガノインにより真珠光沢が付与される。菱形板状で大形、蝶番様構造で互いに連結する。
- コズミン鱗(コズミンりん、英: cosmoid scale)
- 肉鰭類の鱗。デボン紀の魚類には一般的であったが、現生ではラティメリアのみがもつ。骨性の基底板の上に脈管系を伴ったコズミン(英: cosmin)と呼ばれる象牙質層ができ、その表面をエナメロイドが覆う。
- 骨鱗(こつりん、英: bony scale) または 葉状鱗(ようじょうりん、英: leptoid scale)、板状鱗(ばんじょうりん、英: elasmoid scale)
- 硬骨魚類にみられる鱗。真皮に由来する、骨性の円形の薄板。硬い骨質の上層(骨質層)と膠原繊維の下層(繊維板層)からなる。通常は皮膚の薄層がその表面を覆うが、裸出し前端のみが真皮中にあることも多い。前方の鱗が後方のものを覆い、屋根瓦状に配列する。同心円状の線条(隆起線)及び放射状の線条(溝条)がある。
- 円鱗(えんりん、英: cycloid scale)
- 後部の表面(露出面)が滑らかで棘を持たない鱗。硬骨魚類に普通にみられる。形は円形、四角形、卵形など様々。マイワシ(ニシン科)、カタクチイワシ(カタクチイワシ科)、サケ(サケ科)、アユ(キュウリウオ科)、コイ(コイ科)、メダカ(メダカ科)などにみられる。
- 櫛鱗(しつりん、英: ctenoid scale)
- 薄く、後部表面(露出面・自由面)に多数の小棘がある鱗。小棘の発達程度は種や部位によってさまざまである。イットウダイ(イットウダイ科)、ソコダラ類(ソコダラ科)、スズキ(スズキ科)、イサキ(イサキ科)、マダイ(タイ科)、カナガシラ(ホウボウ科)などにみられる。小棘の発達が悪いものを特に有毛鱗(ゆうもうりん、英: ciliated scale)という。
- 側線鱗(そくせんりん、英: lateral line scales)
- 側線上に並んだ鱗。鰓蓋後部上から尾鰭基底までの数を側線鱗数という。
- 有孔側線鱗(ゆうこうそくせんりん、英: pored lateral line scales) または 側線有孔鱗(そくせんゆうこうりん)
- 外部から小孔の見える側線鱗。
- 胸甲(きょうこう、英: corselet)
- 胸鰭基底から後方に伸びる硬い鱗の集まり。マグロ、カツオ類に発達している。
- 隆起線(りゅうきせん、英: ridge) または 環状隆起線(かんじょうりゅうきせん)、環状線(かんじょうせん、英: circuli)、成長線(せいちょうせん)、同心円線(どうしんえんせん)
- 円鱗や櫛鱗の表層に同心円状に並んだ隆起。各隆起線間は魚体の成長速度に比例し、成長の速いときには広く成長帯とよばれ、遅いときには狭くなり休止帯と呼ばれている。大抵年周期で形成される。
- 年輪(ねんりん、英: annulus)
- 年周期で休止帯によって形成される輪紋。休止帯は水温の変化、採餌量の減少、その他の生理的要因によって形成される。
- コンディション輪(コンディションりん、英: condition ring)
- 繁殖期に形成される産卵記号で、体の脂質含量の低下に伴い形成される輪紋。
- 擬年輪(ぎねんりん、英: fake ring, false ring)
- 何らかの生理的要因によって形成される輪紋。
- 溝条(こうじょう、英: groove) または 放射条(ほうしゃじょう、英: radii)
- 円鱗、櫛鱗の外面に放射状に走る小溝。放射状に走るもの以外に前後、上下方向に走るものもあり、これにより鱗は柔軟性を得て魚体の運動を円滑にする。鱗の破損を防いでいる。
- 鱗相(りんそう、英: scale pattern) または 鱗紋(りんもん、英: ground picture)
- 円鱗及び櫛鱗の表面に隆起線と溝条によって形成される紋様。
- 鱗鞘(りんしょう、英: scale sheath)
- 背鰭や臀鰭の基底部を覆う鱗の集合体。鱗で形成された溝状部。
- 鱗板(りんばん、英: bony plate)
- トゲウオやカジカなどの体表を覆う鱗のようにかたくて大きな板状の鱗。有無や数、配列の仕方は分類形質となる。
- 腋鱗(えきりん、英: axillary scale)
- サケ科魚類に独特の、腹鰭基部にある葉状の鱗。
- 稜鱗(りょうりん、英: scute)
- イワシ類の腹縁やアジ類の体側にある肥大した鱗。表面に鋭い稜線を形成している。有無や数は種の識別に用いられる。俗にゼンゴやゼイゴとも呼ばれる。
- 腹部稜鱗(ふくぶりょうりん)
- サッパやコノシロ、イワシなど(ニシン科)の腹中線に並ぶ稜鱗。
- 楯状鱗(じゅんじょうりん)
- アジ類の側線上にある稜鱗。鱗列に沿って稜線を形成する。
呼吸器官
ウィキメディア・コモンズには、 鰓に関連するカテゴリがあります。
 硬骨魚類の鰓の図。Arch = 鰓弓、Raker = 鰓耙、Fillament = 鰓弁 硬骨魚類の鰓の図。Arch = 鰓弓、Raker = 鰓耙、Fillament = 鰓弁
 コイ(コイ科)の鰓。 コイ(コイ科)の鰓。
 ジンベエザメ(テンジクザメ科)の鰓裂(鰓孔)。 ジンベエザメ(テンジクザメ科)の鰓裂(鰓孔)。
 チャイロマルハタ Epinephelus coioides(ハタ科)の鰓耙(口の奥に見える)。 チャイロマルハタ Epinephelus coioides(ハタ科)の鰓耙(口の奥に見える)。
 ヌタウナギ類の呼吸器官。 ヌタウナギ類の呼吸器官。
 迷路器官をもつキノボリウオ(キノボリウオ科)。 迷路器官をもつキノボリウオ(キノボリウオ科)。
- 鰓(えら、英: gill、羅: branchia)
- 呼吸機能や摂食機能、浸透調整機能などをもつ。水中の酸素を取り込み、血中の炭酸ガスを排出する(呼吸機能)。周りの水中の酸素量の85%あまりまで利用できる。
- 水と一緒に飲み込んだ餌を水と分けて咽に送り込む(摂食機能)。
- 内鰓(ないさい、英: internal gill、羅: entobranchia)
- 鰓蓋の中にある鰓。外鰓の対義語。
- 外鰓(がいさい、英: external gill)
- 鰓孔の外に突き出た羽毛状の鰓。ハイギョやポリプテルスの幼生、軟骨魚類の後期胚にある。支持骨を欠く。
- 全鰓(ぜんさい、英: holobranch) または 完全鰓(かんぜんさい)
- 硬骨魚類では鰓隔膜が退縮して鰓弓の部分だけが残り、この前後に鰓隔膜から突出する2列の櫛状となって直接相対し、合する片鰓。
- 鰓蓋(さいがい、えらぶた、羅・英: operculum)
- 硬骨魚類の鰓を外側から覆う保護のための構造。内部はいくつかの骨格要素を持つ。
- 鰓腔(さいこう、さいくう)または 鰓室(さいしつ、英: branchial chamber, gill chamber)
- 鰓の収められている体の腔所。
- 鰓弁(さいべん、英: gill lamella, branchial lamella) または 鰓葉(さいよう)
- 鰓腔の中に詰まっている呼吸器官で、血管が透けて見えるため赤い。ガスや塩類の交換がなされる。鰓弓の外側に並んでいる。表面に多数鰓薄板が並ぶ。
- 鰓蓋鰓(さいがいさい、英: opercular gill)
- 硬骨魚類の鰓蓋の内面に生じ、鰓としての機能を持つ鰓弁。鰓蓋は本来鰓弓に相当する第二咽頭弓から発達したものなので、しばしば機能を持った鰓となり、擬鰓とは異なる。
- 二次鰓弁(にじさいべん、英: secondary lamellae)
- 二次鰓弁の毛細血管の流れる血液の流れは呼吸水の流れと反対である。表面には塩類細胞がある。
- 塩類細胞(えんるいさいぼう、英: choloride cell) または 塩類分泌細胞(えんるいぶんぴつさいぼう、英: choloride secretory cell)
- 硬骨魚類の二次鰓弁の上皮組織にみられる大型の好酸性分泌細胞。ミトコンドリアと滑面小胞体に富み、ナトリウムイオンと塩化物イオンを能動的に排出する。海水魚では塩類細胞基底部は血管に接し、他端はピットを形成して外界に接する。基底部と側面の細胞膜から入り込んだ滑面小胞体が網目状にミトコンドリアを取り巻きながら端部に向かう。海水魚の鰓に多く、淡水魚に少ない。ニジマスやウナギなどの広塩魚を淡水から海水に移すと塩類細胞は急激に数と大きさを増し、ミトコンドリアも増加する。ナトリウム–カリウムATPアーゼ活性が高く、排出の原動力となる。コルチゾルやACTHは塩類細胞とこの酵素活性を増加させる。
- 鰓薄板(さいはくばん、英: second gill lamellae)
- 鰓弁の表面に直立して並んでいる。総面積は広く、平均して体表の表面積の2倍前後ある。
- 鰓裂(さいれつ、英: gill slit, branchial slit, gill cleft, branchial cleft) または 鰓孔(さいこう、えらあな、鰓穴)
- 鰓腔から体外へ呼吸水を出す孔。(脊椎動物全体では、脊椎動物胚の咽頭から外界に開通する裂孔。そのうち魚類では鰓裂に面した前後の壁に多数の鰓弁を生じ呼吸器官となる。)成魚では体外側及び咽頭に面する部位を区別して外鰓裂(外鰓孔)、内鰓裂(内鰓孔)という。鰓裂(咽頭裂)の数は動物により異なり、ヌタウナギ類では6–14対、ヤツメウナギ類では7対(真の眼を含めて名の由来となる)、軟骨魚類では6対、まれに8対、硬骨魚類では5対ある[注釈 1]。
- 鰓膜(さいまく、英: branchiostegal membrane)
- 鰓条骨を繋ぎ、支えている膜。呼吸の際、鰓腔内の水圧を調節する働きがある。左右の鰓膜が峡部で合一するかどうかは魚類の区別に重要な分類形質である。
- 鰓隔膜(さいかくまく、えらかくまく、英: interbranchial septum) または 鰓間隔壁(さいかんかくへき)
- 鰓裂間にある膜(隔壁)。軟骨魚類ではよく発達し長く、その前後の壁に生じる片鰓は完全に隔てられる。硬骨魚類では退縮して短く、その前後の片鰓は全鰓となる。
- 鰓耙(さいは、英: gill raker、羅: cribrum branchiale) または 鰓篩(鰓師、さいし)
- 無顎類を除く魚類にある鰓弓の内側から前方へ向かい並んでいる結節状や繊維状の突起。微細な餌を水から濾し分ける働きがある。
- プランクトン食の種は羽毛状でよく発達し、数が多い。魚食性のものや底生生物を主食とするものは瘤状で数が少ない。鰓耙数や形はよい分類形質となる。
- 鰓歯(さいし、英: gill teeth)
- 鰓耙のうち歯状のもの。
- 片鰓(へんさい、英: hemibranch、羅: hemibranchia) または 半鰓(はんさい)
- 鰓裂の一側(外壁か内壁の鰓弁列)を総称して言う語。
- 擬鰓(偽鰓、ぎさい、英: pseudobranch、羅: pseudobranchia)
- 鰓蓋の内側にあり、支持骨である鰓弓を持たない鰓。軟骨魚類の呼吸孔と硬骨魚類のそれに相当する部位で、顎骨弓と舌骨弓間の第1咽頭裂に当る部位に発達する鰓と系列相同の構造である。軟骨魚類では呼吸孔鰓(こきゅうこうさい、英: spiracular gill)という。動脈血のみが流れ、呼吸作用はない。硬骨魚類では鰓のような外観を保持する場合もあるが、筋肉や結合組織で覆われ、血液を満たし腺状を呈することもある。皮下に埋没することもある。働きは十分に判明していないが、血液の酸素と炭酸ガスの分圧の受容器として働いているという説、ナトリウムの排出に関与しているという説などがある。強力な二酸化炭素排出能力を持つ。その有無は分類形質となる。
- 鰓囊(鰓嚢、さいのう、英: gill sac, gill pouch)
- 円口類において鰓裂の途中が拡張して形成される囊状の器官。その内面に多数の鰓弁を生じ、鰓を形成する。鰓囊は鰓弓骨格に相当する軟骨が複雑に変形した鰓籠により包まれている。
- 咽皮管 (いんひかん、英: pharyngeo-cutaneous duct) または 食道皮管(しょくどうひかん、羅: ductus oesophageo-cutaneus)
- ヌタウナギ類の左体側最後の鰓囊の直後にみられる小管。咽頭内の余分な水分を外に排出するためのもの。鰓裂に由来はするが才能を形成することなく、咽頭と体側外面の間に単純な小管をなす。この外鰓孔は他の外鰓孔より遥かに大きい。ヤツメウナギではこれを欠く。
- 鰓籠(鰓篭、さいろう、英: gill basket, branchial basket)
- ヤツメウナギ類に独特の骨格部分。鰓の流出管を囲み、軟骨からなる複雑な籠状の器官。鰓を保護する。ただしヌタウナギ類では痕跡的。
- 噴水孔(ふんすいこう、英: spiracle) または 呼吸孔(呼吸口、こきゅうこう、英: spiracle)
- 軟骨魚類や基部で分岐した硬骨魚類の一群に見られる、眼の直後、鰓裂の前方に開孔した呼吸器官。サメ類、エイ類、一部のチョウザメ類が持つ。板鰓類では眼の後方にあり、咽頭に通じる小孔。これは本来は顎弓と舌弓の間にある第1鰓裂の痕跡である。エイ類では実際に呼吸孔を通じて呼吸用の水が咽頭に導かれる。
- 上鰓器官(じょうさいきかん、英: suprabranchial organ)
- カムルチーなどの鰓腔上部にみられる、鰓腔や鰓蓋の粘膜が襞状になり、毛細血管が発達して補助的に空気呼吸する器官。発生的起源が迷路器官と異なり、迷路を形成しない。
- 迷路器官(めいろきかん、英: labyrinth) または 迷路状器官(めいろじょうきかん、英: labyrinthform organ)
- キノボリウオなど迷器類(めいきるい、Labyrinthici)に属する魚類における補助的な空気呼吸器官。鰓の上部にあり、舌顎突起で不完全に区画された上鰓腔内に血管の富んだ多数の襞を持つ室を形成している。第1鰓弓の上皮が発達したもので、1–2層の扁平上皮もしくは立方上皮からなり、上皮内血管がみられる。
歯・消化器官
ウィキメディア・コモンズには、 歯に関連するカテゴリがあります。
 コモリザメ Ginglymostoma cirratum(コモリザメ科)の下顎歯。 コモリザメ Ginglymostoma cirratum(コモリザメ科)の下顎歯。
 ネムリブカ(メジロザメ科)の下顎。有作用歯(機能歯)の内側に補充歯が多数ある。 ネムリブカ(メジロザメ科)の下顎。有作用歯(機能歯)の内側に補充歯が多数ある。
- 歯(は、英: tooth)
- 魚類の歯は上下両顎(顎骨)、前鋤骨、舌、口蓋骨、咽頭骨、中篩骨などにある。顎骨以外のものは餌を噛み砕いたり、切り刻んだりするには小さすぎ、口に咥えた餌を逃さないためにあると考えられる。摩耗するとすぐその付近から新たな歯が生え始める。
- 円錐歯(えんすいし、英: conical tooth)
- 断面は丸く、先の尖った錐状の歯。待ち伏せて餌を獲るマエソ、アンコウなどでは大きくて数が多い。高速で泳ぐカツオ、サワラなどでは小さい。
- 臼歯状歯(きゅうしじょうし、英: molar tooth) または 臼歯(きゅうし)
- 噛み合う面が広くて臼のような形をした歯。径が大きくなり、先端が平たい。固い殻を持つ動物を食べるマダイやイシダイなどによく発達する。
- 門歯状歯(もんしじょうし、英: incisor-like tooth)
- 板状で切縁がナイフのように鋭い葉。上顎と下顎で癒合しており噛み切るのに適した歯または、1列に密に並んでいる歯をいう。前者はブダイ科やトラフグ科、後者はイスズミ科で見られる。
- 犬歯状歯(けんしじょうし、英: canine-like tooth) または 犬歯(けんし)
- 長く牙状の歯。円錐歯が肥大かつ彎曲した歯のこと。貪食なミズウオ、タチウオによく発達する。
- 敷石状歯(しきいしじょうし、英: pavemental tooth)
- タイル状の歯
- 鋸歯状歯(きょしじょうし、英: serrated tooth)
- 鋸の歯のような歯。
- 絨毛状歯(じゅうもうじょうし、英: villiform tooth)
- 微細な歯。
- 絨毛状歯帯(じゅうもうじょうしたい)
- 微小な歯が密生した歯帯。
- 上顎歯(じょうがくし)
- 上顎骨に並ぶ歯。
- 前上顎骨歯(ぜんじょうがくこつし)
- 上顎歯のうち前上顎骨に並ぶ歯。
- 主上顎骨歯(しゅじょうがくこつし)
- 上顎歯のうち主上顎骨に並ぶ歯。
- 下顎歯(かがくし) または 歯骨歯(しこつし)
- 下顎骨(歯骨)に並ぶ歯。
- 鋤骨歯(じょこつし)
- 鋤骨に並ぶ歯。
- 口蓋骨歯(こうがいこつし)
- 口蓋骨に並ぶ歯。
- 口蓋歯(こうがいし)
- ヌタウナギ科で口蓋中央にある1本の針状歯。
- 内翼状骨歯(ないよくじょうこつし)
- 内翼状骨に並ぶ歯。
- 副蝶形骨歯(ふくちょうけいこつし)
- 副蝶形骨に並ぶ歯。
- 基舌骨歯(きぜっこつし) または 舌下歯(ぜっかし)
- 基舌骨にある歯。
- 基鰓骨歯(きさいこつし)
- 基鰓骨に並ぶ歯。
- 咽頭歯(いんとうし、英: pharyngeal tooth)
- 咽頭骨に並ぶ歯。食物の咀嚼に使われる。形状は食性と関係している。固い餌を食べるブダイ、ベラ、ウミタナゴ類はよく発達する。
- 上咽頭歯(じょういんとうし、英: upper pharyngeal tooth)
- 上顎の咽頭歯。
- 下咽頭歯(かいんとうし、英: lower pharyngeal tooth)
- 下顎の咽頭歯。
- 歯板歯板(しばん、英: dental plate)
- 大きな歯が互いに癒合したもの。
 円口類のウミヤツメ(ヤツメウナギ科)の歯。 円口類のウミヤツメ(ヤツメウナギ科)の歯。
- 角質歯(かくしつし、英: horny tooth)
- 円口類の口腔およびその周辺や舌にある歯状物。表皮の角質形成物で、脊椎動物の象牙質を持つ真歯とは異なる。円錐形を成し、消耗すると更新される。
- 上唇歯(じょうしんし)
- ヤツメウナギ類の背面にある一群の歯。
- 下唇歯(かしんし)
- ヤツメウナギ類の腹面にある一群の歯。
- 側唇歯(そくしんし) または 内側唇歯(ないそくしんし)
- ヤツメウナギ類の口の左右側面にある歯のこと。
- 周辺歯(しゅうへんし)
- ヤツメウナギ類にある歯。その周りには口縁総状物がある。
- 口歯板(こうしばん、英: lingual lamina)
- ヤツメウナギ類が持つ角質の歯。餌となる生物への吸着や、筋肉を舐めとるといった作用がある。
- 上口歯板(じょうこうしばん、英: supra-pral lamina
- 下より上方にある歯板。ヤツメウナギ類の口歯板の背側。
- 下口歯板(かこうしばん、英: infra-oral lamina)
- 舌より下方にある歯板。ヤツメウナギ類の口歯板の腹側。
- 前舌歯板(ぜんぜっしばん)
- ヤツメウナギ類にある歯板。
- 可倒歯(かとうし、英: flexible tooth)
- 必要に応じて基部が折れ曲がって、内側または奥の方に倒れ、再び元に戻る歯。
- 有作用歯(ゆうさようし、英: functional tooth) または 作用歯(さようし)、機能歯(きのうし)
- 表面上に突出して作用している歯。
- 補充歯(ほじゅうし、英: supplymentary tooth)
- 現在作用している歯の直下または近くにあって、有作用歯が脱落した際発達して代替する歯。
- 外列歯(がいれつし)
- メジナ属で上下顎歯の歯列で摂餌に機能している歯の列。
 魚類の内臓。 魚類の内臓。
- 口腔咽喉囊(口腔咽喉嚢、こうこういんこうのう、こうくういんこうのう)
- ペリカンのように下顎の腹面にできた袋状物。フクロウナギ、ウケグチザラガレイなどの深海魚にみられる。
- 咽喉囊(咽喉嚢、いんこうのう)
- 咽頭にある盲囊。餌となる微生物を集めるのに役立つ。
- 食道(しょくどう、英: esophagus)
- 消化管の一部。
- 食道囊(食道嚢、しょくどうのう、英: esophageal sac)
- 食道にある膨れた袋状物。内面に歯の形をした突起がある。
- 胃(い、英: stomach)
- 消化管の一部。硬骨魚類の中でもコイ類、ダツ類、サンマ類、ベラ類、ブダイ類は胃を欠く。
- 腸(ちょう、英: intestine)
- 消化管の一部。腸の巻き方は近似種間でかなり異なる。
- 螺旋弁(らせんべん、英: spiral valve)
- 腸の内面にある。消化吸収の効果を大きくする。軟骨魚類や原始的な硬骨魚類にある。
- 直腸(ちょくちょう、英: rectum)
- 腸の末端部。
- 腹腔(ふくこう、ふっこう、ふくくう、英: abdominal cavity、羅: cavum abdominis)
- 消化管、鰾、生殖腺、腎臓などの諸器官を包蔵した体腔。
- 腹膜(ふくまく、英: peritoneum)
- 腹腔の内壁および腹腔の中の内部各器官を覆う薄い膜。
- 肝膵臓(かんすいぞう、英: hepatopancreas)
- 膵臓の組織が肝臓内に入り込んだもの。コイ、キス、タイ、ベラ、カワハギなどがもつ。
- 幽門垂(ゆうもんすい、英: pyloric caeca)
- 硬骨魚類の胃と腸の境(幽門部)にある紐状あるいは棍棒状の盲囊。食物の消化と脂肪などの栄養分の蓄積をする働きがある。分類形質となる。
感覚器官とその付属器
 シロカグラ(カグラザメ科)の眼。アーモンド形をしている。 シロカグラ(カグラザメ科)の眼。アーモンド形をしている。
- 眼(目、め、英: eye、羅: oculus)
- 魚類の眼には涙腺がなくレンズが球形で、角膜(かくまく、英: cornea)に接近して位置する。
- 網膜にうまく像を結ぶため、硬骨魚類ではレンズ牽入筋(レンズ収縮筋)の作用でレンズを後方に移動させる。いっぽう軟骨魚類ではレンズは正常な位置で遠くを見るようになっており、近くを見るときはレンズ牽入筋の働きで前方に移動させる。
- 瞳孔(どうこう、英: pupil) または 瞳(ひとみ)
- レンズの前面に虹彩(こうさい、英: iris)があり、その中央部に円形の瞳孔が位置する。多くの硬骨魚類の瞳孔の大きさは明るさによって変化しない。
- 虹彩皮膜(こうさいひまく)
- コチ科魚類(マゴチなど)などにある瞳孔の上部に延長している虹彩の膜。
- 魚類錐体モザイク(ぎょるいすいたいモザイク、英: fish cone mosaic)
- 魚類が持つ、錐体細胞の配置が高い規則性を示す網膜モザイク(もうまくモザイク、英: retinal mosaic)[注釈 2]。例えば、ゼブラフィッシュの網膜では青、赤、緑、紫外線の各波長に感度のピークを持つ4種の錐体細胞が二次元周期的に現れる、正方格子状のパターンがみられる。
- 鎌状突起(れんじょうとっき、かまじょうとっき、英: falciform process、羅: processus falciformis)
- 魚類の眼球にあり、レンズの遠近調節を司る膜状組織。毛様体と同じような働きをする。脈絡膜の一部から突出する眼状体で、やや曲がった先端(ハラー鈴状体)がレンズの中に入っている。鎌状突起内部には平滑筋のレンズ牽入筋があり、この収縮により、レンズは網膜に近づく方向に移動するので遠方に視度を調節できる。
- ハラー鈴状体(ハラーれいじょうたい、羅: campanula Halleri)
- 鎌状突起の曲がった先端にある部分。膨れて丸みがある。
- 眼瞼(がんけん、英: eyelid)
- サメ類にあり、そのためサメの眼はアーモンド形をしている。
- 瞬皮(しゅんひ)
- サメ類の眼の下にある未発達の眼瞼。
- 瞬膜(しゅんまく、英: nictiating membrane、羅: membrana nictitans) または 第三眼瞼(だいさんがんけん、英: tertial palpebra, third eyelid)
- 眼窩内にある角膜の上を覆う1枚の皮膚の皺。異物が目に接近したときに使われる。ネコザメ、トラザメ、オナガザメなど多くの板鰓類がもつ。
- 瞬膜腺(しゅんまくせん、英: gland of nictiating membrane)
- 瞬膜の下、眼の内角に開口する涙腺状の腺。瞬膜が発達している動物に顕著。涙腺と同じように液体を分泌し眼球の表面を潤して角膜と瞬膜との摩擦を緩和する。
- 脂瞼(しけん、英: adipose eyelid)
- 眼を覆う半透明の襞で、脂肪性の眼瞼である。泳ぐ際に頭まわりの水流を変えて水の抵抗を減らす。イワシ、アジ、ボラ、コイ、サバ類などにある。
- 鼻(はな、英: nose、羅: nasus)
- 脊椎動物の嗅受容器。顎口類胚においては神経板の出現前後に体先端の神経板の前端付近で、正中線の左右で表皮の一部が鼻プラコード(はなプラコード、英: nasal placode, olfactory placode、羅: lamina nasalis)としてともに著しく肥厚する。
- 円口類では鼻プラコードは腺性下垂体(下垂体前葉)のプラコードとともに鼻下垂体という正中単一の原基として発し、これに由来する嗅覚器官はさらに落ち込み、鼻管という盲管を形成する。ヌタウナギ類はさらにこれが咽頭に通ずる。板鰓類では鼻は吻下面にあり、口腔と連絡する。他の魚類では両側に鼻孔があるが、口腔と連絡せず、外鼻孔のみを持つ。
- 鼻孔(びこう、英: nostrill, nares, (複:naris)、独: Nasenloch)
- 多くの魚類では外鼻孔のみで内鼻孔はない。
- 内鼻孔(ないびこう、英: internal nostrill, internal nares)
- 口や喉の中に開く鼻孔。ハイギョなどごく一部の原始的な硬骨魚類では内鼻孔があり、鰾で空気呼吸する。
- 外鼻孔(がいびこう、英: external nostril, external nares)
- 吻にあり、外界に開く鼻孔。円形または裂孔状で2対からなる。種によっては1対のものもいる。鼻管の先に開くこともある。
- 2対あるうち前のものを前鼻孔(ぜんびこう)、後ろのものを後鼻孔(こうびこう)といい、においを溶かした水は前鼻孔から入り後鼻孔から出る。
- 鼻腔(びこう、びくう、英: nasal cavity、羅: cavum nasi、独: Nasenhöhle)
- 多くの魚類では表皮下に形成された袋状のくぼみにすぎず口や喉に通じない。
- 嗅板(きゅうばん、英: olfactory lamina)
- 嗅房の中に10枚ほどある、ハート形をした嗅覚器官。表層には嗅覚細胞がある。濁った環境では索餌、索敵などに重要な働きを示す。嗅板の数、配列状態は分類形質となる。
- 嗅房(きゅうぼう、英: olfactory rosettes)
- 嗅板が一定の方式で並んで形成する。
- 鼻管(びかん、英: nasal tube)
- 体表に突き出た小管。先端に鼻孔が開く。
- 鼻弁(びべん)
- 鼻への水の出入りを調整する。
- 嗅窩(きゅうか、英: olfactory pit, nasal sac) または 鼻窩(びか、英: nasal pit、羅: foveanasalis)
- 鼻プラコードの陥入の結果生じた小孔。その一部は嗅上皮を形成する。円口類では無対の嗅窩が正中部に存在する。魚類においては有対の嗅窩がサメ・エイ・ハイギョ類では腹側に、チョウザメ・真骨類では背側にある。内鼻孔類では嗅窩は口陥(こうかん、英: stomadeum)の天蓋に接して口鼻膜(こうびまく、英: oronasal membrane)を形成し、それが開通することにより内鼻孔が生じる。
- 嗅囊(嗅嚢、きゅうのう、英: olfactory sac)
- 円口類の嗅受容器。円口類では鼻孔は1個で頭部の正中線上に開き、ここでは1本の鼻管が鼻囊を作っている。ヤツメウナギ類では鼻囊の中間に嗅囊が開口している。ヌタウナギ類でも同様の嗅囊を持つ。
- 鼻囊(鼻嚢、びのう、英: nasal capsule)
- 円口類の1本の鼻管が作る軟骨性の包被。ヤツメウナギ類では鼻囊が体前端部の背側にある鼻孔に開孔し、先端は脳の下側付近で盲端に終わり、下垂体道(かすいたいどう、独: Hypophysengang)となる。鼻囊の中間には1対の嗅囊が開口し、その後方壁には嗅神経が分布する。ヌタウナギ類では同様の嗅囊を持つが、鼻孔(鼻囊の開口部)は体の前端にあり、下垂体道は食道の前端、前腸に通じている。
- 側線器官(そくせんきかん、英: lateral-line organ) または 側線器(そくせんき)
- 水生脊椎動物(無顎類、魚類、両生類の幼生、水生両生類)の体表にみられる特殊な機械受容器。側線管器と遊離側線器の2種類に分けられる。どちらも基本構造は同じである。感覚細胞とゼリー状のクプラに包まれた有毛細胞とからなる。(仲間、餌、敵などによって生じる)水流や水圧、低周波の音(350Hz以下)、温度変化を受容する。これらの外界の物理的刺激でクプラが曲がると、感覚毛のうちの運動毛を通じて感覚細胞が刺激を受け、興奮する。側線器に生じた刺激は体側では後側線神経(顔面神経に付随している)、頭部では前側線神経(迷走神経に付随している)を通じて延髄に達する。これは脊髄神経によるものではない。
- 側線管器(そくせんかんき) または 側線器(そくせんき)
- 側線管と呼ばれる管や溝が皮下を縦走し、遊離感丘(大孔器)が皮下に埋もれて管状器官の中に収まり、所々で側線孔という小管で外部に開孔する。
- 遊離側線器(ゆうりそくせんき、英: free lateral-line organ)
- 体表に線状もしくは点状に存在し、体の比較的表層に孤立した遊離感丘が頭部から尾部に並ぶ。
- 側線(そくせん、英: lateral line)
- 遊離側線器が外見的に形成するもの。普通、体の中央部の各側に1本あるが、2, 3, 5本など多数の側線を持つものがある。頭部や体側の側線の走り方や側線数は分類上重要な特徴となる。
- 感丘(かんきゅう、英: neuromast, sense hillock) または ニューロマスト(英: neuromast)
- 側線器官の末梢器官。遊離感丘、孔器、管器に分けられる。構造は同一で数個の有毛細胞を中心として有毛細胞と支持細胞からなり、神経は有毛細胞にシナプス構造を形成して接続する。有毛細胞には1本の運動毛と数十本の不動毛がある。
- 遊離感丘(ゆうりかんきゅう) または 大孔器(だいこうき)
- 体表に存在する感丘。
- 孔器(こうき、英: pit organ)
- 孔状になっている感丘。体表の皮膚の中に点在する器官。側線のクプラ、感覚網、感覚細胞のセットが表皮中にある。魚類の孔器の中には1価の陽イオンに反応するものがある。
- 管器(かんき、英: canal organ)
- 皮下に埋没し1本から数本の管状になってその中に存在する感丘。側線のクプラ、感覚網、感覚細胞のセットが皮膚の中に陥入し管の中にある。隣り合う感丘の中間から枝管が出て側線孔により外界に通じている。種によっては側線孔に粘液が詰まっている。
- 有毛細胞(ゆうもうさいぼう、英: stereocilium)
- 側線器官を構成する細胞。有毛細胞には求心性神経と遠心性神経のどちらも分布しているが、後者は前者に対し抑制的に作用する。孔器と管器では2個の有毛細胞が運動毛の局在に対して対をなし、1本の神経線維により支配されている。遊離感丘の場合は必ずしもそうはなっていない。また内耳にも有毛細胞がある。
- 運動毛(うんどうもう、英: kinocilium)
- 有毛細胞にある1本の毛。絨毛構造を示し、運動方向と神経の興奮とに一定の関係があることが明らかになっている。運動毛の周期的運動により膜電位も同様に周期的に変化し、マイクロフォン電位を発生する。2個の細胞が同一繊維により支配されることからマイクロフォン電位は刺激の半分の周期を持つことになり、2倍の周波数を持つ神経電位を生ずることがある。
- 頭部感覚管(とうぶかんかくかん、英: cephalic sensory canal)
- 眼の周囲、下顎、鰓蓋、頭部背面などにある側線器官。
- 眼上管(がんじょうかん)
- 眼の上部にある頭部感覚管。
- 眼下管(がんかかん)
- 眼の後方から下方にある頭部感覚管。
- 鰓蓋下顎管(さいがいかがくかん)
- 鰓蓋前部から下顎に伸びる頭部感覚管。
- 前額把握器(ぜんがくはあくき)
- ギンザメ類が持つ小棘を帯びた軟条突起。交尾の際に鉤着器として用いる。
 電気受容器を持つ弱発電魚のモルミルス属の一種(アロワナ目モルミルス科、エレファントノーズフィッシュに近縁)。 電気受容器を持つ弱発電魚のモルミルス属の一種(アロワナ目モルミルス科、エレファントノーズフィッシュに近縁)。
- 電気受容器(でんきじゅようき、英: electroreceptor)
- 水生動物の体表にあり、体外の電場に対する受容器。魚類では円口類、軟骨魚類(ローレンツィニ器官)、硬骨魚類にみられる。
- ローレンツィニ器官(ローレンツィニきかん、ロレンチーニ器官、英: Lorenzini's organ) または ローレンツィニ瓶器(ローレンツィニびんき、英: Lorenzini's ampulla)、膠質管(こうしつかん、独: Gallertrohr)
- 板鰓類の吻部、頭部側面に分布する皮膚感覚器の一種。体表に無数の開口部を持つ細管が集合して瓶部を作り、中に受容細胞を持つ。元来機械受容器である側線器官の一種で、機械的刺激にも応じる。
- 電気的な受容器としての感度は0.1 μV/cmにも達し、サメやエイでは餌となる魚の運動に伴う活動電位にも反応するため、捕食に利用できるとされる。ゴンズイにも類似した器官があることが知られている。
- (弱電気魚における)電気受容器(でんきじゅようき、英: electroreceptor)
- デンキウナギ目のギュムノートゥス科(Gymnotidae)、アロワナ目のモルミルス科(Mormyridae)とジムナーカス科(Gymnarchidae)の淡水魚は弱電気魚とよばれる。このうち Eigenmannia virescens(アイゲンマニア) は尾部に発電器を持ち250–600 Hzの放電を行って体周辺に電場を作り、伝導体や不導体が近づいた際の電場の乱れを体表にある電気受容器で検出する。
- この電気受容器は側線器の変形したものでローレンツィニ器官とはやや異なる。皮膚の開口部は持たず、皮下に埋没している。放電の頻度は種によって異なるものの、電気の感受性は0.1–100 μV/cmの範囲内にあると考えられている。
- 形状により瓶器(びんき、英: ampullary organ)、瘤状器(こぶじょうき、英: tuberous organ)またはその中間型が区別される。モルミルス科では瓶器、クレノン器、モルミロマストの3種類の受容器があり、それぞれ異なった形態と機能を持つ。この魚の菱脳背側にはきわめてよく発達した電気感覚葉がみられ、3種の受容体からの情報はそれぞれ特定の領域に入力する。
 サヴィ器官を持つゴウシュウシビレエイ(ヤマトシビレエイ科) サヴィ器官を持つゴウシュウシビレエイ(ヤマトシビレエイ科)
- サヴィ器官(サヴィきかん、英: Savi's vesicles)
- シビレエイ類の頭部にある電気器官の周囲の表皮下に多数集まる触感器の一種。完全に閉じた2–3 mmの径を持つ全体を薄膜(クプラ)で包まれた有毛の感丘3個とそれを包む細胞からなる。発電機能を持つほかの魚類には存在しない。
- ウェーバー器官(ウェーバーきかん、英: Weberian apparatus) または ウェーベル氏器官(ウェーベルし・きかん)
- 鰾と内耳を連絡する4小骨片からなる聴覚器官(機械受容器官)。ネズミギス目と骨鰾上目(コイ目、ナマズ目、デンキウナギ目)にみられる。これらは椎骨と肋骨に由来し、四肢動物の耳小骨とは異なる。鰾が受けた圧力波は鰾が接続するウェーバー器官に伝わり、さらにリンパ液に満たされた無対洞、横行管を通って左右の内耳の小囊に伝達される。これによって小囊内の耳石が振動し、小囊斑が刺激され刺激され、鰾が受けた圧力波が知覚される。
鰾
ウィキメディア・コモンズには、 鰾に関連するカテゴリがあります。
 ラッド(コイ科)の有管鰾。 ラッド(コイ科)の有管鰾。
- 鰾(浮き袋、うきぶくろ、英: gas bladder, air bladder, swim bladder)
- 脊椎骨の下、消化管と中腎との間にあり、気体を満たした無対の膜状器官(浮力調節器官)。白色もしくは銀白色の卵形や管状、円錐形の嚢。いろいろな部位から盲囊を出すものもある。隔壁で2–3室に分かれるものもある。気体を出入させて体の比重を調節する。祖先系硬骨魚類の肺に由来し、発生上鰓裂後方部の消化管の膨らみとして生じる。
- ヒラメやカレイでは退化している。そのため急速に水面に引き上げても泳げなくなることはない。(逆に無管鰾をもつ魚類では鰾が膨らみ、泳げなくなる。)
- その他、補助的な呼吸器官、発音器官、聴覚補助器官として働くものもある。
- 有管鰾(ゆうかんひょう、英: physostomous gas bladder) または 有気管鰾(ゆうきかんひょう)
- 気道のある鰾。下等な真骨類がもつ。浮力調節のためのガスの出し入れは短時間でできる。有管鰾を持つ、つまり気道を持つものを開鰾類(かいひょうるい、吸鰾類、きゅうひょうるい、英: physostomi)という。
- 無管鰾(むかんひょう、英: physoclistous gas bladder) または 無気管鰾(むきかんひょう)
- 気道がない鰾、もしくは発生の初期のみにあり変態前後に退縮し、消化管から分離した鰾。進化した真骨魚類が持つ。無管鰾を持つ、つまり気道を持たないものを閉鰾類(へいひょうるい、英: physoclisti)という。
- 発生の初期に食道の背面に鰾が形成され開口の直後までは食道と連絡しているが、発生が進むにつれ食道との連絡は絶たれる。体の比重の調節には時間がかかる。
- 気道(きどう、英: pneumatic duct) または 鰾気管(ひょうきかん、英: pneumatic duct)
- 消化管と鰾を結びつける管状物。真骨類ではチョウザメ、サケ、アユ、コイ、メダカなどの低位グループのみにある。タラ目、スズキ目より高等なグループでは消失している。
- 赤斑(せきはん、英: red spot, red body)
- 鰾に分布する血管によって形成される、1ヶ所または数ヶ所の毛細血管の網目。奇網(きもう、奇驚網、怪網、英: mirabile net、羅: rete mirabile)の一種。
- ガス腺(ガスせん、英: gas gland)
- 血液からの酸素などのガスを出し入れする。鰾内表面の表皮の特化した組織。無管鰾を持つ魚類(閉鰾類)において、食道との連絡を絶たれた後に使用される。ガス腺に入った血液(赤斑)から鰾の中にガスが放出される。
- 赤腺(せきせん、英: red gland)
- 赤斑は閉鰾類ではガス腺と連絡する。赤腺は赤斑とガス腺を合わせて言うもの。二酸化炭素や酸素を分泌する。閉鰾類では卵円腺と合わせ比重の調節に用いられる。
- 卵円腺(らんせんせん、英: ovoid gland) または 卵状体(らんじょうたい)
- 鰾の後背壁にある毛細血管に富む腺。鰾の中のガスを血液に吸収する。閉鰾類では赤腺と合わせ比重の調節に用いられる。
生殖・哺育器官
 ハナザメのクラスパー(交尾器)。 ハナザメのクラスパー(交尾器)。
- 育児囊(育児嚢、いくじのう、英: brood pouch, pouch marsupial、羅: marsupium) または 卵囊(卵嚢、らんのう)
- タツノオトシゴなどヨウジウオ科魚類の雄の体の腹面にみられる溝状または囊状の器官。卵または仔魚を育てる哺育器官である。卵は開口部を過ぎる時雄に受精し、雄は卵を受け取ると育児囊内に胎盤状の組織が発達し、表面が網目状になって一つの卵を収める。卵は40–50日で孵化し、仔魚は臍囊が吸収されるまで育児囊内にいる。その発達程度や位置は分類形質として用いられる。カミソリウオ類では雌にみられる。
- 交尾器(こうびき、英: copulatory organ) または 交尾器官(こうびきかん)、交接器(こうせつき)
- 交尾の時に精液を雌の体内に送り込むための棒状物。サメ類、胎生メダカなどにある。特に板鰓類のものは鰭脚と呼ばれる。
- 鰭脚(ききゃく、クラスパー、英: clasper、羅: pterygopodium)
- サメ・エイ類の雄にある交尾器官。腹鰭の内縁が変化したもので、鰭骨格に由来する軟骨で強化されている。精液輸送のための溝を備え、交尾のとき雌の総排泄孔に挿入する。また、テングヘラザメは雌にも未発達の鰭脚が存在する(雌雄とも機能しない異性の生殖器を持っている)。
- 生殖突起(せいしょくとっき)
- 卵や精子を出す生殖器が単なる穴ではなく肉質の突起状になっている場合を言う。体内受精を行う魚類、例えばカサゴの仲間(フサカサゴ科など)では、成熟すると雄の突起は著しく伸長し、交尾器として働く。
- 泌尿生殖突起(ひにょうせいしょくとっき、英: urinogenital papilla)
- 輸尿管が生殖突起と癒合したもの。交尾の際精液を雌の生殖孔に向け放出する。
- 総排泄孔(そうはいせつこう、英: cloaca) または 総排出腔(そうはいしゅつこう)
- 消化管の終末部で、生殖輸管と輸尿管が同時に開く孔。軟骨魚類にみられる。
- 子宮(しきゅう)
- サメ類の輸卵管の膨大部。
- 胎盤(たいばん、英: placenta)
- 胚組織と母体組織とが密接に接着し、両者間に生理的な物質交換が行われている場合の両者の複合組織。
- 卵黄囊胎盤(卵黄嚢胎盤、らんおうのうたいばん、英: yolk-sac placenta)
- 卵胎生のサメ類、Mustelus および Carcharis の胚の卵黄囊の壁と子宮壁とが接着して形成される。
- 偽胎盤(ぎたいばん)
- 胎生硬骨魚類の一種 Heterandria がもつ。発生が卵胞内で進行し、卵胞壁と心囊とが接着して形成される。
- 鰓胎盤(さいたいばん)
- 胎生硬骨魚類の一種 Jennynsia がもつ。胚が卵黄腔内で発生し、卵巣組織が索状に伸びて鰓裂に侵入し、鰓に接着して形成される。
発光器官
 キュウリエソ属の一種 Maurolicus muelleri(ムネエソ科)とその発光器。 キュウリエソ属の一種 Maurolicus muelleri(ムネエソ科)とその発光器。
- 発光器(はっこうき、英: luminous organ, photogenic organ) または 発光器官(はっこうきかん)
- 魚類には発光する種類が多くみられる。発光を司る器官のことを発光器という。ワニトカゲギス目、ハダカイワシ目などにある。
その他の器官
 タイセイヨウダラ(タラ科)の耳石。 タイセイヨウダラ(タラ科)の耳石。
- 耳石(じせき、英: otolith) または 平衡石(へいこうせき、英: statolith)、聴石(ちょうせき)
- 前庭囊内にある固形物。主として炭酸カルシウムで形成され、平衡を保つのに役立つ。前庭囊は平衡胞の基本構造を保持し、内壁の一部が平行斑となって耳石がこれに接触する。扁平石、礫石、星状石の3種類がある。種によって形が違い、種の特徴となる。また耳石には幅の広い不透明帯と狭い透明帯が同心円状に配列される。これは個体の成長が停滞する時期に有機質の沈着が少なく、透明化することにより形成される。これによってできた紋は年齢や日齢の査定に用いられる。耳石には日周輪(にっしゅうりん、日輪、にちりん、英: daily ring)が示され、仔稚魚の齢が分かるようになった。シログチ(イシモチ)の耳石は特に大きく、名前(石持)の由来となっている。
- 扁平石(へんぺいせき)
- 内耳の球形囊にある耳石。普通は耳石のなかで最大。単に耳石といった場合扁平石を指す。
- 礫石(れきせき)
- 内耳の卵形囊にある耳石。
- 星状石(せいじょうせき)
- 内耳の壺囊にある耳石。
- 球形囊(球形嚢、きゅうけいのう、英: saccule) または 球囊(球嚢、きゅうのう)、小囊(小嚢、しょうのう、羅: sacculus)
- 内耳の膜迷路の一部で、平衡覚受容器。受容細胞は垂直の面上に並ぶ。垂直の運動に対して働く。ヌタウナギでは球形囊と分離せず、ヤツメウナギでは分化する。有毛細胞の大部分は支持細胞から分泌された平衡石膜(平衡砂膜)で覆われ、この平衡石膜に対する重力が常に刺激となっている。静止時でも刺激が加わるため姿勢を保つために重要な役割を果たしている。魚類のこの平衡石は矢(英: sagitta)と呼ばれる。哺乳類の蝸牛は球形囊の基礎部に由来する。
- 卵形囊(卵形嚢、らんけいのう、英: ctricle) または 通囊(通嚢、つうのう、羅: utriculus)
- 球形囊と同じ機能を持ち、構造的にも類似している。受容細胞は水平面に位置する。水平面上の運動に対して働く。ヌタウナギでは卵形囊と分離せず、ヤツメウナギでは分化する。
- 視葉(しよう、英: optic lobe)
- 脳の中央背部にあり、視覚の中枢。
- 嗅葉(きゅうよう、英: olfactory lobe)
- 脳の前部(端脳)にある膨れた部分。嗅覚の中枢。
- 上生体(じょうせいたい、英: epiphysis)
- 嗅葉の後方にある棒状物。光やホルモン分泌に関係している。
- 終脳(しゅうのう、英: end-brain, telencephalon) または 端脳(たんのう)
- 脊椎動物において、前脳の前半分に由来する部位。円口類の終脳は嗅葉として発生する。
- 唇(くちびる、英: lip) または 口唇(こうしん)
- ヤツメウナギのアンモシーテス幼生の口の上下にある襞状構造を上下唇という。また、軟骨魚類の上下顎に付随する襞も「lip」と呼ばれる。
- 上唇(じょうしん、英: upper lip)
- アンモシーテス幼生の口器の背側縁を成す襞状の構造。
- 下唇(かしん、英: lower lip)
- アンモシーテス幼生の口器の腹側縁を成す襞状の構造。
- 皮褶(ひしゅう)
- 皮膚が皺のように盛り上がってできる筋のこと。
- 唇褶(しんしゅう、英: labial fold)
- サメ類の口角部付近にある襞。
- 皮弁(ひべん、英: dermal flap)
- 体表に突き出た皮膚の突出物。舌状、リボン状、分枝状などがある。ミノアンコウの幼魚は全身に著しく長い皮弁が覆う。
- 追星(おいぼし、英: pearl organ) または 真珠器(しんじゅき)
- 繁殖期にできる、表皮が肥厚してできた真珠様の白い突起。多くは雄の前頭部(鰓蓋)、鱗、胸鰭、腹鰭に出現する。種によって出現する場所は異なる。雌に現れる種もあるが、雄のようには発達しない。頭部追星は繁殖期の雄の闘争の際武器として使用される。コイ科、カトストムス科、アユ、一部のスズキ目にみられる
- 真珠器(しんじゅき、英: pearl organ)
- コイ科の追星は表皮細胞が肥厚、突出しその外面を角質化した細胞層が覆う。狭義にはこれのみを真珠器という。
- 接触器(せっしょくき、英: contact organ)
- 河口域に生息するメダカ類 Fundulusの追星。角質層はなく内部に石灰化した硬部を備える。
- 婚姻器(こんいんき、英: nuptial organ)
- 角質層も石灰化した硬部もない。雄性ホルモンの支配下にある二次性徴で、生殖腺を除去した雄では現れない。アユに見られる。
 コバンザメ(コバンザメ科)の背面。吸盤があり、これは第1背鰭が変化したもの。 コバンザメ(コバンザメ科)の背面。吸盤があり、これは第1背鰭が変化したもの。
 ヌマチチブ(ハゼ科)の腹面にある吸盤。腹鰭が変化したもの。2つある腹鰭の間は膜蓋でつながっている。 ヌマチチブ(ハゼ科)の腹面にある吸盤。腹鰭が変化したもの。2つある腹鰭の間は膜蓋でつながっている。
- 吸盤(きゅうばん、英: sucker)
- 他物に吸着する装置。ハゼ科、ウバウオ科にある腹面の吸盤は腹鰭の変形物、コバンザメの吸盤は第1背鰭の変形物である。
- 膜蓋(まくがい)
- 吸盤状になっているハゼ類の腹鰭の左右にある棘を連結させている膜のこと。
 エスカをもつカエルアンコウ(カエルアンコウ科)。これで餌となる生物を誘き寄せる。 エスカをもつカエルアンコウ(カエルアンコウ科)。これで餌となる生物を誘き寄せる。
- 擬餌状体(ぎじじょうたい、イリシウム、英: illicium) または エスカ(英: esca、疑似餌、カエルアンコウ科の擬餌状体)、ルアー(英: lure)
- アンコウ類などの吻上にある誘引突起の先端にある膨らんだ部分。これを動かして餌となる生物を引き寄せる。 ここから発光物質を放出する種もある。タイコウボウダルマのルアーには眼のような一対の小黒斑と付属肢に見える糸状物を備え、エビに似せている。
- 誘引突起(ゆういんとっき) または 第1背鰭棘(だいいちせびれきょく)、吻上棘(ふんじょうきょく、カエルアンコウ科の誘引突起)
- アンコウ類などの頭の背面にある突起。これを振って先端の擬餌状体が生きているように見せかける。背鰭第1棘が変化したもの。タイコウボウダルマなども持つ。
- 皮膚腺(ひふせん、英: dermal gland) または 皮腺(ひせん)
- 脊椎動物の皮膚線は魚類、両生類、哺乳類によく発達する。魚類の皮膚腺は単細胞性または多細胞性の粘液腺(英: mucous gland)で粘液を分泌する。また一部は鰭棘に毒腺があり、深海魚の発光腺にも皮膚腺に属するものがある。
- 粘液孔(ねんえきこう)
- ヌタウナギ類の体表にある。
- 粘液囊(粘液嚢、ねんえきのう、英: mucilage sac)
- ヌタウナギ類において表皮中の単細胞粘液腺のほかに体側の各体節ごとに存在する多細胞性の粘液腺。糸細胞(しさいぼう、英: thread cell)という特異な細胞からなり、細胞内の長さ数cm、幅1–3 μmの粘液糸(ねんえきし、英: mucous thread)を大量の粘液とともに放出して水中に粘液網を貼り、捕食や防御に使われる。
- 塩類腺(えんるいせん、英: salt gland) または 直腸腺(ちょくちょうせん)
- 海産の軟骨魚類などにある塩化ナトリウム分泌腺。その他爬虫類、鳥類が持つ。塩化ナトリウムを海水よりも濃い溶液として排出する。軟骨魚類では直腸腺という。シーラカンスは硬骨魚類だが、直腸腺を持つ。分泌細胞はミトコンドリアに富み、構造は哺乳類の尿細管細胞に似ている。
- 尾部神経分泌系(びぶしんけいぶんぴつけい、英: caudal neurosecretory system)
- 魚類の脊椎骨末端付近に分布する神経分泌細胞の細胞体と軸索、軸索の末梢部を含む尾部下垂体系の総称。マンボウ科とヨウジウオ科を除く硬骨魚類と板鰓類に存在し、全頭類には知られていない。板鰓類やチョウザメ類には分泌細胞はあっても尾部下垂体は存在しない。
- 背刀(はいとう、英: dorsal blade)
- テンガンムネエソ、ムネエソモドキなどムネエソ類の背鰭基部前方にある。
- 体輪(たいりん)
- ヨウジウオ科の体はかたい体輪で覆われている。
- 軀幹輪(躯幹輪、くかんりん)
- 軀幹部を覆う体輪。
- 尾輪(びりん)
- 尾部を覆う体輪。
- 凹窩(おうか)
- サメ類の尾鰭の付け根の凹みのこと。
- 欠刻(けっこく、英: notch)
- 切れ込み。特にサメの尾鰭上葉先端付近にある切れ込みを指す。
 発電器官をもつアイゲンマニア Eigenmannia virescens(ステルノピュグス科) 発電器官をもつアイゲンマニア Eigenmannia virescens(ステルノピュグス科)
- 発電器官(はつでんきかん、英: electric organ) または 電気器官(でんききかん)
- 発電魚の発電力を持つ器官。筋肉または軸索が分化して形成される。シビレエイやアイゲンマニアなどでは頭の両側、デンキウナギやデンキナマズでは軀幹部、ガンギエイでは尾部にある。短い時間間隔で自らの周囲に電場を形成し、その電場の乱れを感知して餌や外敵の存在を知るのに利用される。
- デンキウナギでは終板電位に相応するシナプス電位とこれに続く活動電位の発生によって神経支配面は外側が-70 mVになり他の面では静止電位が保たれ外側が+80 mVなので1つの電函を挟んで150 mVの電位が発生する。デンキウナギの電気柱は5000–6000の電函が同方向に直列的に繋がっているので、活動電位が同期して発生すると600–860 Vの電圧を発生させる。なお活動電位の持続は神経や筋と同じくミリ秒のオーダーである。
- シナプス電位だけで活動電位を発生しないものもおり、電気柱を構成する電函の数も数十の場合から数千にもわたるので、全起電力は1–3 Vのものから、ミシマオコゼ類約5 V、シビレエイ類では30–80 V、デンキナマズでは400–450 V、デンキウナギでは600–860 Vにも及び、まちまちである。デンキナマズのものは皮膚の腺組織が分化したもので、電函の起電力の方向が逆である(頭部がマイナス、尾部がプラス)。
- サックス器官(サックスきかん、英: Sachs organ)
- デンキウナギが600–860 Vの起電力の発電器官とは別に、体の後部に持っている起電力の弱い発電器官。構成する電函の起電力は主器官の起電力と同じ(百数十mV)だが、各電函の間隔が大きい。方向探知機の発信機としての機能を持つと考えられている。
- 電函(でんかん、英: electroplaque) または 電気板(でんきばん、英: electroplate)
- 発電器官の最小単位のシンシチウム。多数集まり電気柱を形成。電函の一側面は神経支配を密に受けており、この神経伝達物質はアセチルコリンである。
- 電気柱(でんきちゅう)
- 電函が積み重なってできたもの。多数の電気柱が平行に並んで1個の発電器官を形成する。
- 棘(とげ、英: spine)
- 体表に突き出た骨質隆起で、先端が尖ったもの。
- 鰭棘(ききょく)
- 鰭の棘。ネコザメの背鰭の棘や、エイの尾棘など。
- 上膊棘(じょうはくきょく、英: humeral spine)
- 胸鰭基底の上方付近で体表に突き出た棘。アンコウの上膊棘は多尖頭、メダマアンコウの上膊棘は2尖頭、キアンコウの上膊棘は単尖頭である(尖頭も参照)。
- 擬棘(ぎきょく)
- タラ科魚類の鰭の棘。
- 額棘(がっきょく)
- カサゴ類などの頭部背面にある棘のうち、前頭骨隆起(両眼の間にみられる隆起)の後端にある棘。
- 頸棘(頚棘、けいきょく)
- カサゴ類などの頭部にある棘。頭部背面の頭頂骨隆起の後方に位置する。
- 耳棘(じきょく)
- カサゴ類などの頭部にある棘。
- 鼻棘(びきょく)
- カサゴ類などの頭部にある棘。
- 翼棘(よくきょく)
- ガンギエイ類などの胸鰭にある棘。
- 眼前棘(がんぜんきょく)
- カサゴ類などの頭部にある棘。
- 眼上棘(がんじょうきょく)
- カサゴ類などの頭部にある棘。
- 眼後棘(がんこうきょく)
- カサゴ類などの頭部にある棘。
- 眼部棘(がんぶきょく)
- ガンギエイ類などの眼付近にある棘。
- 項部棘(こうぶきょく)
- ガンギエイ類などの項部にある棘。
- 肩帯部棘(けんたいぶきょく)
- ガンギエイ類などの肩帯部にある棘。
- 尾部棘(びぶきょく)
- ガンギエイ類の腰帯後部から第1背鰭までの背中線上にある棘。
- 眼窩後棘(がんかこうきょく)
- カサゴ類などの頭部にある棘。
- 上擬鎖骨棘(じょうぎさこつきょく)
- カサゴ類などの上擬鎖骨付近にある棘。
- 上後側頭棘(じょうこうそくとうきょく)
- カサゴ類などの頭部にある棘。
- 下後側頭棘(かこうそくとうきょく)
- カサゴ類などの頭部にある棘。
- 後側頭骨棘(こうそくとうこつきょく)
- ムネエソ類などの頭部にある棘。
- 前鰓蓋骨棘(ぜんさいがいこつきょく)
- カサゴ類、ムネエソ類などの頭部にある棘。
- 前部腹縁棘(ぜんぶふくえんきょく)
- ムネエソ類などの腹部にある棘。
- 後部腹縁棘(こうぶふくえんきょく)
- ムネエソ類などの腹部にある棘。
- 離棘(りきょく)
- 臀鰭の前方にある小棘。アジ類には2本の離棘がみられる。
 アメリカアカエイ Dasyatis Americana(アカエイ科)。尾には尾棘が見える。この尾棘には毒腺がある。 アメリカアカエイ Dasyatis Americana(アカエイ科)。尾には尾棘が見える。この尾棘には毒腺がある。
- 尾棘(びきょく、英: tail-spine)
- エイ類の尾部に生えている棘。尾棘の両端は鋸歯縁となり毒腺を持つ。一般にアカエイ、トビエイ、ツバクロエイなど尾部背中線上にあるものをいう。ガンギエイにあるものは尾部棘と区別する。
- 出鰓血管(しゅっさいけっかん、英: efferent branchial vessel)
- 鰓で酸素を得た動脈血を鰓から心臓または全身に送る血管。解剖学上は鰓静脈(さいじょうみゃく、英: branchial vein)。
- 側静脈(そくじょうみゃく、英: lateral vein、羅: vena laterails)
- 魚類のみにある体側筋中から静脈血を心臓へ導く左右1対の血管。その走行中に腹鰭からくる腸骨静脈及び胸鰭からの鎖骨下静脈を受けキュヴィエ管に合し、それが心臓の静脈洞に開く。
骨格
ウィキメディア・コモンズには、 骨格に関連するカテゴリがあります。
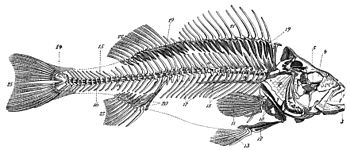 魚類の骨格。1.前上顎骨(独: Zwischenkiefer)、 魚類の骨格。1.前上顎骨(独: Zwischenkiefer)、
2.主上顎骨(独: Oberkiefer)、
3.下顎骨(独: Unterkiefer)(特に歯骨)、
4.眼窩(独: Augenhöhle)、
5.眼下骨(独: untere Augenknochen)、
6.前鰓蓋骨(独: Vordeckel)、
7.下鰓蓋骨(独: Unterdeckel)、
8.間鰓蓋骨(独: Zwischendeckel)、
9.主鰓蓋骨(独: Deckel)、
10.肩帯(独: Schultergürtels)、
11.胸鰭(独: Brustflosse)、
12.腰帯(独: Beckenknochen)、
13.腹鰭(独: Bauchflosse)、
14.脊柱(独: Wirbelsäule)、
15.神経棘(棘突起)(独: obere Dornfortsätze)、
16.血管棘(独: untere Dornfortsätze)、
17.肋骨(独: Rippen)、
18.上肋骨(独: Fleischgräten)、
19.神経間棘(担鰭骨)(独: obere Flossenträger)、
20.血管間棘(担鰭骨)(独: untere Flossenträger)、
21.背鰭の棘条(独: Stacheln der Rückenflosse)(第1背鰭、棘条背鰭)、
22.背鰭の軟条(独: Strahlen der Rückenflosse)(第2背鰭、軟条背鰭)、
23.臀鰭(独: Afterflosse)、
24.下尾骨(独: senkrechte Schwanzplatte)、
25.尾鰭(独: Schwanzflosse)
 魚類の骨格。青色が担鰭骨(背鰭担鰭骨、臀鰭担鰭骨、肩帯、腰帯) 魚類の骨格。青色が担鰭骨(背鰭担鰭骨、臀鰭担鰭骨、肩帯、腰帯)
 オウギチョウチョウウオ(チョウチョウウオ科)の骨格。 オウギチョウチョウウオ(チョウチョウウオ科)の骨格。
- 頭骨(とうこつ)
- 頭の骨。
- 頭蓋骨(ずがいこつ、とうがいこつ、英: skull)
- 脳を保護する骨。頭骨のうち両顎部、舌部、鰓部の骨を除いたもの。特に分類学上重要である。
- 篩骨(師骨、しこつ、英: ethmoid (bone))
- 鼻殻域にある頭蓋骨の一部。
- 上篩骨(上師骨、じょうしこつ、英: supraethmoid (bone))
- 鼻殻域にある頭蓋骨の一部。
- 側篩骨(側師骨、そくしこつ、英: lateral ethmoid (bone))
- 鼻殻域にある頭蓋骨の一部。
- 鋤骨(じょこつ、英: vomer) または 前鋤骨(ぜんじょこつ、英: prevomer)
- 鼻殻域にある頭蓋骨の一部。頭蓋骨の最前端にある不対の骨。これに歯があれば口蓋部の正中線上に露出する。歯の有無は分類に用いられることもある。
- 鼻骨(びこつ、英: nasal (bone))
- 鼻殻域にある頭蓋骨の一部。小さな膜骨。
- 前頭骨(ぜんとうこつ、英: frontal (bone))
- 眼の周辺にある頭蓋骨の一部。
- 眼窩蝶形骨(がんかちょうけいこつ、英: orbitoshenoid (bone))
- 眼の周辺にある頭蓋骨の一部。眼球の上方、眼窩の後部にある。スズキ目などの進化した魚類では欠き、ニシン科、コイ科、キンメダイ科魚類など下等なグループが持つ。ウナギ目もこれを欠く。
- 翼蝶形骨(よくちょうけいこつ、英: pterosphenoid (bone))
- 眼の周辺にある頭蓋骨の一部。
- 基蝶形骨(きちょうけいこつ)
- 眼の周辺にある頭蓋骨の一部。眼窩の後下方にある不対の小さい骨。タラ目魚類のような底生魚では欠く。
- 鞏膜骨(きょうまくこつ)
- 眼の周辺にある頭蓋骨の一部。
- 蝶耳骨(ちょうじこつ、英: autosphenoid (bone))
- 耳殻域にある頭蓋骨の一部。
- 翼耳骨(よくじこつ、英: autopterotic (bone))
- 耳殻域にある頭蓋骨の一部。
- 前耳骨(ぜんじこつ、英: prootic (bone))
- 耳殻域にある頭蓋骨の一部。
- 上耳骨(じょうじこつ、英: epiotic (bone))
- 耳殻域にある頭蓋骨の一部。
- 外後頭骨(がいこうとうこつ、英: exoccipital (bone))
- 耳殻域にある頭蓋骨の一部。
- 上後頭骨(じょうこうとうこつ、英: supraoccipital (bone))
- 耳殻域にある頭蓋骨の一部。頭蓋骨のうち、後部背方の正中線にある骨。
- 頭頂骨(とうちょうこつ)
- 耳殻域にある頭蓋骨の一部。頭蓋骨のうち後部前方にある骨。
- 間在骨(かんざいこつ、英: opisthotic (bone), intercalar (bone))
- 耳殻域にある頭蓋骨の一部。ウナギ目はこれを欠く。
- 基後頭骨(きこうとうこつ、英: basioccipital (bone))
- 床域にある頭蓋骨の一部。
- 副蝶形骨(ふくちょうけいこつ、英: parasphenoid (bone))
- 床域にある頭蓋骨の一部。
- 眼窩(がんか)
- 眼を収めている骨の凹所。
- 涙骨(るいこつ、英: lachrymal (bone)) または 眼前骨(がんぜんこつ、英: preotbital bone)
- 眼の周囲に並ぶ数個の小骨のうち最前のもの。
- 眼下骨(がんかこつ、英: suborbital (bone), infraobital (bone))
- 眼の下方に一列に並んだ歯のうち涙骨を除いたものの総称。前方のものから第1眼下骨、第2眼下骨というように呼ぶ。
- 眼下骨棚(がんかこつほう、英: suborbital stay)
- 第2眼下骨が後下方へのびたもの。カサゴ目に特有。
- 上神経棘(じょうしんけいきょく、英: )
- 背鰭から前方の頭骨にかけて筋肉の背中線にある小さな骨。胴長の種ほど多い。
- 上顎骨(じょうがくこつ、英: upper jaw)
- 上顎を構成する骨。前上顎骨と主上顎骨の総称。
- 前上顎骨(ぜんじょうがくこつ、英: premaxillary)
- 上顎を構成する骨の一つ。
- 主上顎骨(しゅじょうがくこつ、英: maxillary)
- 上顎を構成する骨の一つ。
- 下顎骨(かがくこつ、英: lower jaw)
- 下顎を構成する骨。硬骨魚類では歯骨、角骨、後関節骨の総称。
- 歯骨(しこつ、英: dentary)
- 下顎を構成する骨の一つ。
- 角骨(かくこつ、英: angular)
- 下顎を構成する骨の一つ。
- 後関節骨(こうかんせつこつ、英: retroarticular)
- 下顎を構成する骨の一つ。
- 上側頭骨(じょうそくとうこつ、英: supratemporal bone)
- 肩帯の最上方にある小骨。
- 後側頭骨(こうそくとうこつ、英: posttemporal bone)
- 肩帯の上部を占め、頭骨に連なる。
- 喉板(こうばん、英: gular plate)
- 左下の下顎間にある板状の骨。下等な硬骨魚類にある。
- 懸垂骨(けんすいこつ)
- 口蓋諸骨と鰓蓋諸骨の総称。
- 口蓋諸骨(こうがいしょこつ) または (狭義の)懸垂骨(けんすいこつ、英: suspensorium)
- 口蓋を構成する骨の総称。鰓蓋諸骨と合わせて懸垂骨という。
- 口蓋骨(こうがいこつ、英: paratine)
- 上顎の内側にある骨。口蓋にある1対の骨。ここに歯があれば前鋤骨の外後方の両側に露出し、重要な分類形質となる。
- 内翼状骨(ないよくじょうこつ、英: endopterygoid)
- 眼の内下面にある膜状の骨。
- 外翼状骨(がいよくじょうこつ、英: ectopterygoid)
- 眼の内下面にある棒状の骨。
- 後翼状骨(こうよくじょうこつ、英: metapterygoid)
- 眼の内下面で内翼状骨の後方にある骨。
- 方骨(ほうこつ、英: quadrate)
- 舌顎骨(ぜつがくこつ、英: hyomandibular)
- 接続骨(せつぞくこつ、英: symplectic)
- 鰓蓋諸骨(さいがいしょこつ)
- 鰓蓋を構成する4つの骨の総称。口蓋諸骨と合わせて懸垂骨という。
- 前鰓蓋骨(ぜんさいがいこつ、英: preopercle)
- 鰓蓋を形成する4つの骨のうち、背前方にある骨。
- 主鰓蓋骨(しゅさいがいこつ、英: opercle)
- 鰓蓋を形成する4つの骨のうち、背後方にある骨。
- 間鰓蓋骨(かんさいがいこつ、英: interopercle)
- 鰓蓋を形成する4つの骨のうち、前腹方にある骨。
- 下鰓蓋骨(かさいがいこつ、英: subopercle)
- 鰓蓋を形成する4つの骨のうち、後腹方にある骨。
- 舌弓(ぜっきゅう、英: hyoid arch) または 舌弧(ぜっこ)
- 角舌骨、間舌骨、下舌骨、上舌骨、尾舌骨、鰓条骨からなる。
- 咽舌骨(いんぜっこつ) または 基舌骨(きぜっこつ、英: basihyal (bone))
- 舌を支える骨。1個。鰓弓の最前部に位置する。側面から見ると中央部が最も高く、次第に低くなり端部が最も低い。
- 角舌骨(かくぜっこつ、英: ceratohyal)
- 鰓条骨を支える骨。1対。
- 間舌骨(かんぜっこつ、英: interhyal)
- 鰓条骨を支える骨を懸垂骨の接続骨付近に関節させる。1対。
- 下舌骨(かぜっこつ、英: hypohyal)
- 鰓条骨を支える骨。1対。
- 上舌骨(じょうぜっこつ、英: epihyal)
- 鰓条骨を支える骨。1対。
- 尾舌骨(びぜっこつ、英: urophyal)
- 咽舌骨の下方に懸垂する骨。1個。
- 鰓条骨(さいじょうこつ、英: branchostegal (rays))
- 鰓蓋の後下縁につながる膜骨。鰓膜を支持し、鰓孔の開閉、すなわち呼吸水の保持と排出に役立つ。4–12対ある。
- 鰓弓(さいきゅう、英: gill arch, branchial arch) または 鰓弧(さいこ)
- 鰓耙と鰓を支えている骨。5対あり、基鰓骨、下鰓骨、角鰓骨、上鰓骨、咽鰓骨、下咽頭骨から構成される。上下咽頭骨はそれぞれ咽頭歯を備えている
- 基鰓骨(きさいこつ、英: basibranchial bone)
- 鰓を支持する骨格で、口内の床面中央部に前から後へ1列に並ぶ(中軸部を直走する)。通常3個の骨からなる。
- 下鰓骨(かさいこつ、英: hypobranchial bone)
- 角鰓骨(かくさいこつ、英: ceratobranchial bone)
- 咽頭の床部を構成する。
- 上鰓骨(じょうさいこつ、英: epibranchial bone)
- 咽頭の天井部を構成する。
- 咽鰓骨(いんさいこつ、英: pharyngobranchial bone)
- 咽頭の天井部を構成する。第4-5番目の咽鰓骨が癒合して上咽頭骨(じょういんとうこつ、英: upper pharyngeal bone)になる。
- 上咽頭骨(じょういんとうこつ、英: upper pharyngeal bone)
- 第4–5番目の咽鰓骨が癒合したもの。
- 下咽頭骨(かいんとうこつ、英: lower pharyngeal bone)
- 第5鰓弓の下半の2つの骨が癒合したもの。
- 脊椎骨(せきついこつ、英: vertebrae)
- 頭骨後端から尾端まで体の中軸にある骨。多くの硬骨魚類においてよく骨化している。脊椎骨の数は種によってほぼ一定の範囲内にあり、よい分類形質となる。
- 椎体(ついたい、英: centrum)
- 脊椎を形成し体軸に並んだ個々の骨。ハイギョ、チョウザメなどは発達していない。腹椎と尾椎に分かれる。
- 腹椎(ふくつい、英: abdominal vertebrae) または 腹椎骨(ふくついこつ)
- 軀幹部の脊椎骨。原則、肋骨がついている。第1腹椎は基後頭骨(頭蓋骨後端基部にある)に関節する。スズキ目では普通10本ある。
- 尾椎(びつい、英: caudal vertebrae) または 尾椎骨(びついこつ)
- 尾部の脊椎骨。尾椎の最後端の1–3本は尾骨となる。スズキ目では普通14–15本ある。
- 神経棘(しんけいきょく) または 棘突起(きょくとっき)、棘状突起(きょくじょうとっき)
- 脊椎骨の上部(背側)にある突起。
- 横突起(おうとっき)
- 脊椎骨の下部(腹側)にある突起。尾椎のものは血管棘となる。
- 血管棘(けっかんきょく)
- 尾椎において横突起が下方で癒合し形成されたもの。癒合した基部では尾動脈を収める血道弓門となる。
- ウェーバー骨片(ウェーバーこっぺん、英: Weberian ossicle) または ウェーバー小骨(ウェーバーしょうこつ)
- 第1–4椎体から分かれた4つの小骨片からなり、内耳と鰾を結びつけて聴覚の能力を高める。ウェーバー器官を構成する。ネズミギス目と骨鰾上目のみにある。
- 三脚骨(さんきゃくこつ、英: tripus)
- ウェーバー器官を構成する骨片の一つ。鰾に接続する。
- 挿入骨(そうにゅうこつ、英: intercalarium)
- ウェーバー器官を構成する骨片の一つ。
- 舟状骨(しゅうじょうこつ、英: scaphium)
- ウェーバー器官を構成する骨片の一つ。
- 結骨(けっこつ、英: claustrum)
- ウェーバー器官を構成する骨片の一つ。無対洞に接続する。
- 肋骨(ろっこつ、ろくこつ、英: rib、羅: costa)
- 腹部の椎体の横突起から側下方に出る棒状の骨。内臓を囲む体壁を維持する。
- 背肋(はいろく、英: dorsal rib) または 上肋(じょうろく、独: obere Rippe)
- 筋系を背腹に分ける水平筋中隔中に生じる肋骨。軟骨魚類や四肢動物が持つ。
- 腹肋(ふくろく、英: ventral rib, pleural rib) または 下肋(かろく、独: untere Rippe)
- 血道突起が血道弓を成さず、左右に開いたまま体腔壁直下に伸びたもの。硬骨魚類が持つ。魚類の中には背肋を併用するものもあり、ハイギョの後頭骨にある頭肋(とうろく)も下肋である。
- 上肋骨(じょうろっこつ、英: epipleural rib)
- 魚類がもつ軸上筋間中にある肋骨。
- 肩帯(けんたい、英: shoukder girdle)
- 胸鰭を支持する骨。四肢動物の前肢の骨の由来となる。軟骨魚類のものは肩胛軟骨、烏口軟骨、基底軟骨、輻射軟骨の4つからなり、単純である。硬骨魚類のものは上擬鎖骨、擬鎖骨、後擬鎖骨、烏口骨(中烏口骨)、射出骨、肩胛骨からなり、複雑である。
- 硬骨魚類のもの
-
- 擬鎖骨(ぎさこつ)
- 肩帯を構成する骨。
- 上擬鎖骨(じょうぎさこつ)
- 肩帯を構成する骨。
- 後擬鎖骨(こうぎさこつ)
- 肩帯を構成する骨。ウナギ目はこれを欠く。
- 烏口骨(うこうこつ、英: coracoid) または 烏喙骨(うかいこつ)
- 肩帯を構成する骨。
- 中烏口骨(ちゅううこうこつ、英: mesocoracoid) または 中烏喙骨(ちゅううかいこつ)
- 肩帯を構成する骨。ニシン、サケ類には見られるが、スズキ、カジカ類には見られない。
- 射出骨(しゃしゅつこつ、英: actinost) または 輻射骨(ふくしゃこつ)
- 肩帯を構成する骨。
- 肩胛骨(肩甲骨、けんこうこつ)
- 肩帯を構成する骨。
- 軟骨魚類のもの
-
- 肩胛軟骨(肩甲軟骨、けんこうなんこつ)
- 軟骨魚類の肩帯を構成する軟骨。
- 烏口軟骨(うこうなんこつ)
- 軟骨魚類の肩帯を構成する軟骨。
- 基底軟骨(きていなんこつ、英: basalia)
- 軟骨魚類の肩帯を構成する軟骨。対鰭の近心部に1個から数個ある。
- 輻射軟骨(ふくしゃなんこつ、英: radialia)
- 軟骨魚類の肩帯を構成する軟骨。対鰭の遠心部に多数ある。
- 腰帯(ようたい、英: pelvic girdle)
- 腹鰭を支持する骨。四肢動物の後肢の骨の由来となる。一本の腰骨からなる。ニシン目など下等な硬骨魚類では体の後方に位置するが、スズキ目など高等な魚類ではそれが前方に移動した位置となり、肩帯に接する。
- 腰骨(ようこつ、英: pelvic bone)
- 腰帯を構成する骨。
- 担鰭骨(たんきこつ、英: pterygiophore, fin suspensorium)
- 背鰭や臀鰭の基部にあり、背鰭や臀鰭を支持する骨。肩帯や腰帯を含めていうこともある。科レベルでは差異はなく近縁種を比較する材料とはならない。
- 近担鰭骨(きんたんきこつ、英: proximal pterygiophore) または 近位担鰭骨(きんいたんきこつ)
- 近心側の担鰭骨。
- 神経間棘(しんけいかんきょく)
- 背鰭の(近)担鰭骨。脊椎骨の神経棘の間に位置する。
- 血管間棘(けっかんかんきょく)
- 臀鰭の(近)担鰭骨。脊椎骨の血管棘の間に位置する。
- 遠担鰭骨(えんたんきこつ、英: distal pterygiophore) または 遠位端鰭骨(えんいたんきこつ)
- 遠心側の担鰭骨。
- 基鰭軟骨(ききなんこつ)
- 軟骨魚類の不対鰭を支持する軟骨。
- 近基鰭軟骨(きんききなんこつ、英: basal segment)
- 最も近位の基鰭軟骨。
- 間基鰭軟骨(かんききなんこつ、英: median segment)
- 近基鰭軟骨と遠基鰭軟骨の間の基鰭軟骨。
- 遠基鰭軟骨(えんききなんこつ、英: distal segment)
- 最も遠位の基鰭軟骨。
- 尾部棒状骨(びぶぼうじょうこつ)
- 最後の尾椎と第1尾鰭椎。
- 下尾骨(かびこつ、英: caudal skeleton) または 尾鰭担鰭骨(びきたんきこつ、おびれたんきこつ)
- 尾鰭を支持する骨。尾鰭を支えるために扇状に広がっている。脊椎骨の血管棘が肥大して形成された。
筋肉
ウィキメディア・コモンズには、 筋肉に関連するカテゴリがあります。
- 体側筋(たいそくきん、英: lateral mussle)
- 軀幹部、尾部の尾側によく発達している筋肉。体側筋の収縮により遊泳の推進力が生産される。運動量の多いものほどミオグロビンが多く含まれ、その量により白色(白身魚)、桃色、赤色(赤身魚)を呈する。
- 血合筋(ちあいきん、英: dark mussle) または 血合肉(ちあいにく)
- 体側筋のうち濃赤褐色をした筋肉。多くは体表付近にある。毛細血管に富み、運動力の大きい魚類はよく発達する。マグロ類は体表付近のみならず中心部にもある。
- 筋節(きんせつ、英: myomere, myotome)
- 頭索動物および魚類や両生類(幼生と有尾類成体)の体幹に前後軸に沿って分節的に並ぶ筋。体側筋に規則的に配列される一つ一つの筋肉の束。W状に1列に並び、それぞれは筋節中隔(きんせつちゅうかく、英: myoseptum)で仕切られている。1個の筋節には1本の脊髄神経が対応するように配置されている。
体形
 琵琶形のビワアンコウ Ceratias holboelli(ミツクリエナガチョウチンアンコウ科) 琵琶形のビワアンコウ Ceratias holboelli(ミツクリエナガチョウチンアンコウ科)
 ヒシダイ Antigonia capros(ヒシダイ科)。菱形をしている。 ヒシダイ Antigonia capros(ヒシダイ科)。菱形をしている。
魚の体形は体色とともに分類形質として重要であり、生息場、生活様式によってさまざまなものがある。魚類の代表的な形状は紡錘形(ぼうすいけい)である。カエルアンコウやダンゴウオなどは球形と表現される。コショウダイ、ゴマモンガラ、コウベダルマガレイなどは卵形、ハナビラウオなどは卵円形(らんえんけい)、ギンガメアジの幼魚、アオスジモンガラなどは長卵形(ちょうらんけい)をしている。また、ヒトスジタマガシラなどは半円形(はんえんけい)、クロダイ、チョウチョウコショウダイなどは楕円形(だえんけい)、タマガシラ、サザナミウシノシタなどは長楕円形(ちょうだえんけい)と表現される。シワイカナゴ、ボラ、ヒメコダイ、ボウズカクレウオなどのように円筒形(えんとうけい)をしているものやウナギのレプトケファルス幼生のようにリボン形をしているもの、ヒシダイ、メイタガレイなどのように菱形(りょうけい、ひしがた)をしているものやシラウオなどのように楔形(くさびがた、英: wedge-shaped)のものもいる。
以下のような用語も用いられる。
- 側扁形(そくへんけい、英: compressiform) または 側扁(そくへん)
- 体高が高く、体幅が狭いもの。遊泳魚に多い。「側扁する」のように使う。エンゼルフィッシュやヒラメ、スイ、マダイやイシダイなど。特に、ツバメウオやイトヒキアジのように体高が体長の半分を上回る場合、強く側扁すると表現される。
- 縦扁形(じゅうへんけい、英: depressiform) または 縦扁(じゅうへん)
- 体高が低く、体幅が広いもの。底生魚に多い。「縦扁する」のように使う。エイやアンコウ、ウミテングなど。
- 体盤(たいばん、英: disc)
- 躯幹部、頭部などが一体となり縦扁した形。エイやアンコウなど。
- 延長形(えんちょうけい) または 延長(えんちょう)
- 体長が体高の8倍以上あるもの。「延長する」のように使う。アシロ目など。
模様
- 縦帯(じゅうたい、英: longitudinal band) または 縦縞(たてじま)、縦班(じゅうはん)、縦線(たてせん)
- 体の長軸に沿って走る色帯(縞模様・斑紋・細い縞模様)。頭を上、尾部を下にして魚体を立たせた場合の表現。ヌノサラシなど。
- 横帯(おうたい、英: cross band) または 横縞(よこじま)、横斑(おうはん)、横線(よこせん)
- 体の長軸方向と直角の方向の色帯(縞模様・斑紋・細い縞模様)。頭を上、尾部を下にして魚体を立たせた場合の表現。ブリモドキなど。
- 斜帯(しゃたい) または 斜走帯(しゃそうたい)、斜縞(しゃじま)
- 斜めに走る色帯(縞模様)。
- 放射帯(ほうしゃたい)
- 1点から放射状に走る帯。
- 鞍状斑(あんじょうはん)
- 背から振り分けた形の斑紋。馬の鞍のようであることから。
- 円斑(えんはん) または 円形紋(えんけいもん)
- 円形の斑紋。
- 眼状斑(がんじょうはん、英: eye-spot) または 眼状紋(がんじょうもん)
- 淡色に縁どられた円形の暗色斑紋。
- 虫食い状斑(むしくいじょうはん)
- 虫が食った後のような複雑な斑紋。
- パーマーク(英: parr mark)
- サケ科魚類のみにみられ、幼魚期に体側にある斑紋。成魚になると消えるものが多いが、河川残留型のものは斑紋が消えない。
その他の特徴
- 鋸歯縁(きょしえん、英: serrated margin)
- 歯や棘などの縁が鋸歯となること。
- 切縁(せつえん、英: cutting edge)
- 縁辺がナイフで切断したような形をしているときに用いる。
- 櫛状(しつじょう)
- 櫛の歯のような形態のこと。櫛鱗、櫛状歯などという使い方をする。
- 尖頭(せんとう、英: cusp) または 尖頭状(せんとうじょう)
- 歯の先端やサメ類の鱗の後端などが山状または針状に尖ること。出っ張りが1つのとき単尖頭、2つのとき2尖頭、3つのとき3尖頭という。サメの鱗には単尖頭、三尖頭、四尖頭、五尖頭、多尖頭状などがみられる。
- 截形(せつけい、英: truncate)
- 縁辺がまっすぐとなった形。尾鰭や胸鰭などの後縁の形を表現する際に用いる。
- 湾入形(わんにゅうけい、わんにゅうがた、英: emerginate)
- 縁辺が丸く引っ込んだ形。
- 楔形(くさびがた、英: wedge-shaped)
- 縁部が突出した形のこと。尾鰭後縁やまれに体形などに用いる。
各部の位置
- 端位(たんい)
- 口の位置。口が頭の前端に開くこと。
- 亜端位(あたんい)
- 口の位置。端位より幾分後方に口が開く状態。
- 上位(じょうい)
- 口の位置。口が頭の前端より上に開くこと。アンコウ類など。
- 下位(かい)
- 口の位置。口が頭の前端より下に開くこと。ギンザメなど。
- 斜位(しゃい)
- 口の位置。マトウダイ科、ヒシダイなど。
- 胸位(きょうい)
- 腹鰭の位置を表す用語。胸鰭の下方近くに腹鰭が位置する場合に用いる。
- 喉位(こうい)
- 腹鰭の位置を表す用語。腹鰭が胸鰭下方より前方に始まっている場合に用いる。
- 腹位(ふくい)
- 腹鰭の位置を表す用語。腹鰭基底の下部から肛門までの間に腹鰭が位置する場合に用いる。
- 正中線(せいちゅうせん) または 体軸(たいじく)
- 魚体の中心部を縦方向(頭の前端から尾部)に貫く線のこと。
- 背中線(はいちゅうせん)
- 魚体を上から見た時、背面中央を頭から尾に引いた線。魚を観察する際の基準として用いる。
- 腹中線(ふくちゅうせん)
- 魚体を下から見た時、腹面中央を頭から尾に引いた線。魚を観察する際の基準として用いる。
- 体側(たいそく)
- 体の側面のこと。
測定部位
- 全長(ぜんちょう、TL、英: total length)
- 体の最前端から尾鰭の後端までの長さ。尾鰭はすぼめてその後端までを測定する。
- 体長(たいちょう、BL、英: body length) または 標準体長(ひょうじゅんたいちょう、SL、英: standard length)
- 吻端(必ず上顎)から尾鰭基底(脊椎骨末端(=下尾骨後端)まで)の長さ。尾鰭を左右にまげて折れ曲がるところを目印にして測定する。
- 尾叉長(びさちょう、FL、英: fork length) または 尾叉体長(びさたいちょう)
- 吻の前端から尾鰭の湾入部までの直線距離。
- 体高(たいこう、英: body depth)
- 軀幹部で背腹方向に最も高い部分の直線距離。鰭を含めない。
- 体幅(たいふく、英: body width)
- 軀幹部で左右方向に最も幅広い部位の幅。
- 頭長(とうちょう、英: head length)
- 吻端から主鰓蓋骨の後端までの直線距離。ただし、サメでは最後の鰓裂の後端までの長さで、ヤツメウナギ類では第1鰓孔の後端までの長さ。
- 頭高(とうこう、英: head depth)
- 後頭部の中央における高さ。後頭部から頭の下端または胸部までの高さ。
- 頭幅(とうふく、英: head width)
- 鰓蓋を閉めた状態においての頭部の最大幅。
- 体盤長(体板長、たいばんちょう)
- アンコウ類では頭長と同じ。エイ類では吻端から胸鰭の末端までの長さ。
- 体盤幅(体板幅、たいばんはば、たいばんふく)
- アンコウ類では頭幅と同じ。エイ類では両胸鰭間の最大幅。
- 尾部長(びぶちょう、英: tail length)
- 肛門から尾鰭起部までの直線距離。
- 尾柄長(びへいちょう)
- 臀鰭の最後の軟条の基底から尾鰭中央基底までの長さ。
- 尾柄高(びへいこう、英: depth of caudal peduncle)
- 尾鰭の付け根付近で最も細い部位の長さ。
- 基底長(きていちょう、英: length of fin base)
- 背鰭、臀鰭、尾鰭などの基部両端間の直線距離。
- 背鰭基底長(せびれきていちょう)
- 背鰭第1鰭条の基底から背鰭最終軟条基底までの長さ。背鰭が2–3基の場合、それぞれの基底長を計測することが多い。
- 臀鰭基底長(しりびれきていちょう)
- 臀鰭第1鰭条の基底から臀鰭最終軟条基底なでの長さ。臀鰭が2基の場合、それぞれの基底長を計測することが多い。
- 鰭の長さ(ひれのながさ、英: fin length)
- 不対鰭においては最も長い鰭条の始部から先端までの長さ。胸鰭では最上の基部から最長軟条の先端までの直線距離。腹鰭では起部から最長軟条の先端までの直線距離。
- 胸鰭長(むなびれちょう)
- 最上または最前の腹鰭鰭条の基底から胸鰭の先端までの長さ。
- 腹鰭長(はらびれちょう)
- 最外側または最前の腹鰭鰭条の基底から腹鰭の先端までの長さ。
- 尾鰭長(おびれちょう)
- 尾鰭の基底中央からその先端までの長さ。
- 尾鰭下葉長(おびれかようちょう)
- 尾鰭基底から尾鰭下葉の後端までの長さ。
- 軀幹長(躯幹長、くかんちょう、英: trunk length) または 胴長(どうちょう)
- 鰓蓋の後端から肛門(総排泄孔)までの直線距離。
- 吻長(ふんちょう、英: snout length)
- 吻端から眼の前縁までの距離。
- 口前吻長(こうぜんふんちょう)
- アカエイ類などの口から体の前端までの長さ。
- 口幅(こうふく)
- 口裂の最大幅。
- 上顎長(じょうがくちょう)
- 前上顎骨の前端から主上顎骨の後端までの長さ。
- 下顎長(かがくちょう)
- 下顎の先端からその最後端までの長さ。
- 肛門前長(こうもんぜんちょう、英: preanal length) または 前肛門長(ぜんこうもんちょう)、頭胴長(とうどうちょう)
- 体の前端から肛門の中央までの直線距離。アナゴやタチウオなどの尾部が細く切れやすいものに用いられる。
- 背鰭前長(せびれぜんちょう) または 前背鰭長(ぜんはいきちょう、英: predorsal length)
- 吻端(体の前端)から背鰭起部までの直線距離。
- 腹鰭前長(はらびれぜんちょう)
- 吻端から腹鰭前端までの距離。
- 胸鰭前長(むなびれぜんちょう)
- 吻端から胸鰭第1条基底までの距離。
- 臀鰭前長(しりびれぜんちょう)
- 吻端から臀鰭起部までの距離。
- 尾部前長(びぶぜんちょう)
- 体の前端から肛門までの距離。アカエイ類などで用いる。
- 腹鰭肛門間隔(はらびれこうもんかんかく)
- 腹鰭基底前端から肛門中央までの距離。
- 尾鰭上葉前長(おびれじょうようぜんちょう)
- 軟骨魚類における吻端から尾鰭上葉起部までの長さ。
- 尾鰭下葉前長(おびれかようぜんちょう)
- 軟骨魚類における吻端から尾鰭下葉起部までの長さ。
- 鰓域長(さいいきちょう)
- 第1鰓孔から最後の鰓孔までの長さ
- 両眼間隔幅(りょうがんかんかくふく、りょうがんかんかくはば)
- 両眼間の最短の幅。
- 眼後長(がんこうちょう)
- 眼の後縁から主鰓蓋骨の後端までの長さ。
- 眼下幅(がんかはば、がんかふく)
- 眼窩の下縁から涙骨または眼下骨の下縁までの最小幅。
- 眼径(がんけい)
- 眼の水平的な最大直径。
- 眼窩径(がんかけい)
- 眼を収めている骨の凹所の水平径。
計数形質
略称は中坊徹次『日本産魚類検索 全種の同定 第三版 I』による。
- 鰭条数(きじょうすう)
- 鰭条数は種によっておおむね一定で同定形質となり、鰭式で表す。学術的には棘条をローマ数字、軟条をアラビア数字で表示する。同一の鰭で棘条部と軟条部が分離している場合は「‐」(ハイフン)、連続しているときは「, 」(コンマ)その数字を分ける。また、胸鰭や腹鰭で不分枝軟条数を小文字のローマ数字ⅰ、ⅱ…)、分枝軟条数をアラビア数字で表すこともある。
- 鰭式(きしき)
- 鰓の構造は鰭式で表される。鰭式では背鰭をD(dorsal fin)、第1背鰭をFD(first dorsal fin)、臀鰭をA(anal fin)、胸鰭をP1(pectoral fin)、腹鰭をP2(pelvic fin)、尾鰭をC(caudal fin)などの略号を用いて表す(例:マダイの鰭式:D XII,10 ; A III,8 ; P1 15 ; P2 I,5)。
- 小離鰭は小文字のローマ数字(ⅰ、ⅱ…)と表されることが多い。
- 側線鱗数(そくせんりんすう、LL)
- 側線上の1縦列の全鱗数で肩帯に接する鱗(鰓孔の上端付近)から下尾骨後端までの数。
- 側線孔数(そくせんこうすう、LP)
- 側線上の1縦列の全孔数で、側線鱗数と同様に数える。鱗がない、もしくは退化的なものに用いる。
- 肛門前側線孔数(こうもんぜんそくせんこうすう、PLp)
- 鰓孔の上端付近から肛門直上までの側線孔数。ウナギ目でよく用いられる。頭部が延長しているウナギ目魚類では始部は鰓孔よりかなり前にある。
- 縦列鱗数(じゅうれつりんすう、LR)
- 側線のない種や側線を中断するものに用いられ、側線鱗数と同様に数える。
- 側線有孔鱗数(そくせんゆうこうりんすう)
- 有孔鱗と無孔鱗が側線列上にある場合の有孔鱗数を数える。始点と終点は側線鱗数の数え方と同じ。
- 横列鱗数(おうれつりんすう、TR)
- 一般的には、第1背鰭から小鱗も含めて後下方へ腹部正中線まで走る1横列の鱗数。
- 側線上方横列鱗数(そくせんじょうほうおうれつりんすう、TRa) または 側線上横列鱗数(そくせんじょうおうれつりんすう)
- 第1背鰭の起部から小鱗を含めて後下方に走る側線鱗の一つ手前までの1横列鱗数。側線鱗を含めない。
- 背鰭棘条部中央下側線上方横列鱗数(せびれきょくじょうぶちゅうおうかそくせんじょうほうおうれつりんすう、TRac)
- 背鰭棘条部中央の背鰭基底から小鱗も含め、後下方へ側線鱗の一つ手前の鱗まで走る1横列鱗数。
- 側線下方横列鱗数(そくせんかほうおうれつりんすう、TRb) または 側線下横列鱗数(そくせんかおうれつりんすう)
- 臀鰭起部から小鱗を含めて前上方に走る側線鱗の一つ手前までの1横列鱗数。側線鱗を含めない。
- 背鰭前方鱗数(せびれぜんぽうりんすう、Pred.S)
- 背鰭基部から後頭部に至る正中線上の全ての鱗の数。
- 頬鱗数(きょうりんすう)
- 眼から前鰓蓋骨の隅角にいたる線を横切る鱗列数。
- 尾柄周囲鱗列数(びへいしゅういりんれつすう)
- 尾柄の最も低い部分の1円周鱗数。チョウザメ科の硬鱗は背側のものをDS、体側のものをLS、腹側のものをVSと表記する。
- 筋節数(きんせつすう)
- 筋節の数。幼魚において重要な分類形質。
- 鰓耙数(さいはすう、GR)
- 鰓耙数は鰓耙の上枝と下枝に分けて数え、+の符号で区別する。両枝の中央にあるものは下枝に含めて数える。
- 鰓条骨数(さいじょうこつすう、BR)
- 鰓条骨(鰓蓋の下方にあり、角舌骨、上舌骨に付着し鰓膜を支えている肋骨状の骨)の数。
- 脊椎骨数(せきついこつすう)
- 脊椎骨(背骨を構成する骨)の数。最前の椎体から最後の椎体まで数え、尾鰭を支えるために扇状に広がっている下尾骨を1個として含める。腹椎と尾椎は+の符号で区別する。通常軟X線撮影にて計数される。
- 幽門垂数(ゆうもんすいすう、PC)
- 幽門垂は大部分の硬骨魚類に見られ、種により数が安定しているため、幽門垂数はよい分類形質となる。
- 咽頭歯数(いんとうしすう、Ph)
- コイ科などでは咽頭歯がよく発達し、その数と列がよい分類形質となる。計数法は外側から列ごとに咽頭歯の数を数える。
- 体輪数(たいりんすう)
- タツノオトシゴやヨウジウオがもつ体輪の数。
生態
繁殖・発生
 ウナギ(ウナギ科)の生活環(ライフサイクル)。降河回遊を行う。 ウナギ(ウナギ科)の生活環(ライフサイクル)。降河回遊を行う。
- 生活史(せいかつし、英: life history)
- 生まれてから死亡するまでの個体または種の生活過程。
- 成魚(せいぎょ、英: adult)
- よく成長し、繁殖能力を備えた魚。最初の成熟に達してから、それ以降の魚。
- 未成魚(みせいぎょ)
- 鱗が出来上がってから最初の成熟に達するまでの発育段階。
- 若魚(わかうお、英: young, adolescent)
- 種の特徴は明らかであるが、体形に少しの差異があるもの。
- 幼魚(ようぎょ、英: fry)
- 孵化してから成魚になるまでの一般的な名称。仔魚、稚魚、未成魚ほど厳格に扱われていない。
- 稚魚(ちぎょ、英: juvenile)
- 全ての鰭の条数が成魚の鰭の条数と同数になってから鱗が出来上がるまでの期間。体の各部比、色彩、生態など成魚とかなり異なる。
- 仔稚魚(しちぎょ)
- 仔魚と稚魚を合わせていう言葉。
 卵黄囊を持つサケ類(サケ科)の仔魚。 卵黄囊を持つサケ類(サケ科)の仔魚。
- 仔魚(しぎょ、英: larvae)
- 孵化してから各鰭の鰭条数が成魚と同数になるまでの幼生。
 アナゴ(アナゴ科)のレプトケファルス幼生。 アナゴ(アナゴ科)のレプトケファルス幼生。
- レプトケファルス幼生(レプトケファルスようせい、英: leptocephalus)または 葉形仔魚(ようけいしぎょ)、葉形幼生(ようけいようせい)
- ウナギ目、カライワシ目などの幼生。体は大型で、リボン状(柳の葉のような形)をし、透明である。レプトケファルス幼生の体には細胞外マトリックスであるグリコサミノグリカン(英: glycosaminodlycan、GAG)という粘液多糖類が大量に蓄積される。レプトケファルス、レプトセファルス幼生、レプトセファルスなどの表記もみられる。
- トリクティス幼生(トリクティスようせい、英: tholichthys)
- チョウチョウウオ類(チョウチョウウオ科、クロホシマンジュウダイ科)の幼生。頭が大きく、皮膚が固く、突起がある。トリクティス、トリクチス幼生、トリクチスなどの表記もみられる。
- アンモシーテス幼生(アンモシーテスようせい、アンモシーテス、英: ammocoetes)
- ヤツメウナギ目の幼生。成魚との形態的差異が激しい。口は漏斗状で、眼は皮下に埋没している。口には歯がなく、口内の繊毛運動で微細有機物(プランクトンやデトリタス)を濾過摂食する。河川の泥中に棲み、生育域は一つの河川系に限られる。成長すると体は短くなり、頭巾のような唇は吸盤に、口内の繊毛は歯に変わり、眼も現れる。2基の背鰭が完成し、尾鰭と分離する。種によって幼生期は3–7年であり、日本産のものは3.5–4年程度である。
- 変態(へんたい)
- 幼魚期には成魚と著しく異なった形態をしているものが、急に成魚に類似した姿に変化すること。ウナギ目、カレイ目、マンボウ科などで有名。再演的変態と後発的変態がある。
- 再演的変態(さいえんてきへんたい)
- その動物の先祖の形態(系統発生の途中の一時期の形態)が幼時に一度あらわれてのちに成体の形に変わるもの。サヨリ科、ダツ科などのように成魚の著しく長い吻は後期仔魚期の初めごろまでは全くなく、後仔魚期から稚魚期にかけて急に発達する。またカレイ目の魚も後期仔魚期までは左右対称であるが、後期仔魚期の終わりに急に眼が片側に回り、非対称となる。
- 後発的変態(こうはつてきへんたい)
- 幼時の形態が先祖の形態とまったく関係のないもの。チョウチョウウオ科、カノコウオ科の魚は幼時には頭部の骨格の一部が棘状に伸びている。また、レプトケファルス幼生もこれに当る。金平糖のような棘があるマンボウ科の幼魚もこれである。
- 卵生(らんせい、英: oviparity)
- 卵巣卵が成熟して体外に放出されたのちただちに体外に排出され発生する現象。
- 単卵生(たんらんせい)
- サメ類の卵生の一タイプ。受精卵が輸卵管に移動するとすぐに2個ずつ産卵され、これが繰り返される。軟骨魚類ではネコザメ目、多くのトラザメ類、一部のテンジクザメ目、ギンザメ類、ガンギエイ類に見られる。
- 複卵生(ふくらんせい)
- サメ類の卵生の一タイプ。受精卵がしばらくの間子宮にとどまり、胚がある程度成長してから産卵されるもの。ナガサキトラザメなどにみられる。
 単為生殖するウチワシュモクザメ(シュモクザメ科)。胎盤による胎生種である。 単為生殖するウチワシュモクザメ(シュモクザメ科)。胎盤による胎生種である。
- 胎生(たいせい、英: viviparity)
- 胚が母体内で胎盤などによって栄養を取り、よく発達して出産すること。
- 卵黄依存型胎生(らんおういそんがたたいせい、らんおういぞんがたたいせい)
- サメ類の胎生(卵胎生)の一タイプ。自分の卵黄だけで成長するもの。真正のもの(アブラツノザメなど)と偶発胎生(ジンベエザメなど)がある。
- 母体依存型胎生(ぼたいいそんがたたいせい、ぼたいいぞんがたたいせい)
- サメ類の胎生(卵胎生)の一タイプ。母親から栄養をもらい成長するもの。無精卵や自分自身の卵黄を食べるもの(ホホジロザメなど。ときに兄弟を食べるもの(シロワニなど)もいる)と子宮壁から分泌される子宮ミルクを栄養とするもの(アカエイなど)、胎盤により栄養を受け取るもの(シュモクザメ類など)の3つがある。
- 卵胎生(らんたいせい、英: ovoviviparity)
- 雌の体内で卵が孵化し、稚魚となって体外へ出てくるもの。胎生との区別は曖昧であり、近年は卵胎生と胎生は厳密に区別するべきものではないと考えられ始めている。浮性卵を生む魚などに比べ産卵数が少ない。ウミタナゴ、メバルなど。
- 性転換(せいてんかん、英: sex reversal, sex change)
- 一生のうち性が逆転し、雄から雌、または雌から雄に変化する現象。また、卵巣部と精巣部を持つ雌雄同体のもので、一方の性が機能したあともう一方の性が機能することもいう。ハタ科、タイ科、ベラ科、チョウチョウウオ科、スズメダイ科、キンチャクダイ科などの魚にみられる。
- 雌性先熟(しせいせんじゅく、英: protogyny)
- 先に雌として働いた後、雄に性転換する現象。ソードテール、ミズウオ、ハタ、タウナギ、カンムリベラ、キュウセンなど。
- IP(始相、英: Initial Phase)
- 色彩斑紋が地味な個体。ベラ科、ブダイ科などの、雌性先熟をする個体とそうでない個体が存在する魚種に対して使われる用語。IPは雄と雌両方が存在する。
- TP (終相、英: Ter-minal Phase)
- 色彩斑紋が派手な個体。ベラ科、ブダイ科などの、雌性先熟をする個体とそうでない個体が存在する魚種に対して使われる用語。TPは雄のみである。
- 一次雄(いちじおす)
- 孵化時から雄の個体。性転換して雄となった二次雄が存在する雌性先熟の種のうち、もとから雄である個体を区別したいときに使われる用語。通常、雌と外見が同じである。
- 二次雄(にじおす)
- 雌が性転換して雄となった個体を指す。もとから雄である一次雄が存在する雌性先熟の種のうち、性転換して個体を区別したいときに使われる用語。雌と外見が異なる場合が多い。
- 雄性先熟(ゆうせいせんじゅく、英: protandry)
- 初めは雄として働き、後に性転換して雌になる現象。クロダイ、コチなど。
- 単為生殖(たんいせいしょく、英: parthenogenesis) または 処女生殖(しょじょせいしょく)
- 雌が雄と関係なしに単独で新個体を生ずる生殖法。ギンブナのある個体群やアマゾンモリーなどでは集団の中に雄がおらず、雌の産んだ卵が近縁種の精子により賦活され発生し、それが雌の個体となる(偽受精)。このことをジノゲネシス(英: gynogenesis)という。またウチワシュモクザメは一定の環境下で単為生殖をおこなう。
- 一時性徴(いちじせいちょう)
- 雌雄の生殖腺や生殖器にみられる性徴。
- 二次性徴(にじせいちょう、英: secondary sexual character)
- 成熟により性ホルモンの作用で生殖腺以外の部位の大きさ、構造、色合いなど(体、頭部、鰭などの大きさや形、眼の位置、発光器の形と位置、育児囊の有無、色彩)が雌雄で異なること。
- 托卵(たくらん、英: brood parasitism)
- 他個体に卵を托し、育てさせること。鳥類のカッコウ科などが有名だが、近年魚類でも知られるようになった。托卵する相手の個体を仮親(かりおや、英: host)という。タンガニーカ湖に棲むカッコウナマズは口内保育するシクリッド類に托卵し、その稚魚は口内で仮親の稚魚を食べ成長する(種間托卵、英: intraspecific brood parasitism)。
- 擬似産卵(ぎじさんらん、英: pseudospawning)
- 主に魚類で見られる、実際に卵を放出ことのない産卵行動。スズメダイやカワスズメなどにみられる。繁殖期以外にも観察されたり、雄同士のペアでも行われたりする。繁殖とは直接関係なく個体間での社会的関係の調整機能などを果たしているとも考えられているが、その適応的意味はほとんどわかっていない。
- 偽受精(ぎじゅせい、英: false fertilization)
- 卵に精子が侵入するにも拘らず精核と卵核が合一するに至らないまま退化し、しかも以後の発生が卵核だけの関与によって進行する現象。魚類でもギンブナなど数例が確認されている。ギンブナは関東地方などでは全個体が雌のことがあるが、これは近縁種のキンブナやニゴロブナの精子によって雌性発生するためである。
- 求愛行動(きゅうあいこうどう、英: courtship behavior)
- 産卵に際し、番を組む、または組もうとする相手に対してとる行動。
- 婚姻色(こんいんしょく、英: naptial color)
- 産卵期(サケ科では特に河川遡上期)に現れる独特の色彩。産卵期に最も顕著になり、これを過ぎると消失する。雄に現れることが多い。二次性徴で体色に変化が現れる。サケ科では海洋生活をしていたころの銀色が黒ずみ、赤や橙系統の色が加わって鮮やかになる。
- 同時に生殖腺に筋肉から栄養が行くため、肉の脂や蛋白質、赤身は消えてゆく。雄は鼻曲がりになる。
- 鼻曲がり(はなまがり)
- サケ科魚類などで雄の成魚の両顎が曲がった状態になること。
- 銀化(銀毛、ぎんけ、スモルト、英: silvering, smolt)
- 孵化した稚魚が成長していく段階で体色がゆっくりと銀色に変わっていくこと。サケ科では尾鰭と背鰭の先端が黒ずみ、他の各鰭も透明度を増し、体全体も白っぽくなる。体つきは細くなり、痩せた魚のようになる。淡水域の河川から塩水の海へ出ていく準備。0歳の初秋のころ起こる。甲状腺ホルモンが関与していると考えられる。ウナギも銀化変態し海での生活に適応する。腹が銀色となった銀ウナギはカウンターシェイディングにより見つかりにくい。
- スモルト(英: smolt)は銀化個体(ぎんけこたい)を指す場合もある。
- 擬似銀化(ぎじぎんけ、シュードスモルト、英: pesudo smolt)
- 雨が豊富で、上流の森が保水力があるため水量が多く、個々の縄張りを持つ必要のない場合や、生息する水域が広く行動する範囲が広い場合、体色が銀色となること。起こる理由は縄張りを持たなくなるからである。
- 矮雄(わいゆう、英: dwarf male)
- 雌に比べ体が著しく小さい雄。体制も極度に退化し、雌に寄生している場合が多い。魚類ではアンコウなどにみられる。ミツマタヤリウオの雌は50 cm程度で黒いが、雄は3 cmほどの大きさで成熟し、白っぽく、歯も消化管も退化する。
- 寄生雄(きせいゆう、英: parasitic male)
- 同一種の雌に寄生する雄。矮雄である。例えばビワアンコウでは、雌は体長120 cmにも達するが、雄は10 cm程度しかなく、自由遊泳期を経てメスの体表に吸着し寄生生活に入り、雌の体に完全に癒着する。この雄は呼吸器系、消化器系が退化し、精巣だけが発達する。雌との間には血管の連絡がある。
- サテライト(英: satellite) または 衛星雄(えいせいおす)
- 縄張り防衛や求愛などを行わず、他の雄の繁殖努力に便乗してもしくはその隙をついて繁殖を行う個体。ただし研究者によって定義が異なる。M.Grossによるとスニーカーが成長し、雌擬態によって繁殖中のペアに近づくようになったものを指す。
- スニーカー(英: sneaker)
- 水草の陰にひそみ、産卵放精中の番を見つけると突進して放精する小型雄(M. Grossによる定義、サテライトと区別せず用いることも多い)。
- スニーキング(英: sneaking)
- 水草の陰に潜んでいて、産卵放精中のペアを見つけると突進して放精すること(M. Grossによる定義)。
- ストリーキング(英: streaking)
- 小型雄が雌とペア産卵している大形雄に突進する行動。集団で繁殖する魚種またはハレムを形成する魚種にみられる。
 口内保育しているアフリカン・シクリッドの一種(カワスズメ科)。 口内保育しているアフリカン・シクリッドの一種(カワスズメ科)。
- 口内保育(口内哺育、こうないほいく、マウスブルーディング、英: mouthbrooding) または マウスブリーディング(英: mouth breeding)
- 卵もしくは仔魚を口の中に入れて特定の時期まで親魚がこれを保護すること。
- 口内保育魚(口内哺育魚、マウスブルーダー、英: mouthbrooder) または マウスブリーダー(英: mouth breeder)
- 口内保育する魚。雌は数週間の間絶食に耐える。カワスズメ科、テンジクダイ科、ハマギギ科、アロワナ科、クサウオ科、アゴアマダイ科、キノボリウオ科、オスフロネムス科、タイワンドジョウ科など10科以上。カワスズメ科は主に雌(雌雄交替で口内保育する種もある)が、テンジクダイ科(ネンブツダイ、キンセンイシモチなど)では雄が口内保育する。
- 生殖隔離(せいしょくかくり)
- 集団間で交雑が起こらないこと。
- 基質産卵魚(きしつさんらんぎょ)
- 石や流木、水草の広い葉などの上に卵を産み付ける魚。エンゼルフィッシュなど。
 泡巣でのベタ(オスフロネムス科)の産卵。 泡巣でのベタ(オスフロネムス科)の産卵。
- 泡巣(あわす、英: bubble nest)
- ベタなどアナバンディ類の発情した雄が水面に口を使って盛り上げる細かい泡。雄は雌をこの巣の下に誘い込み、自分の体をU字型に曲げて雌を抱きしめ、産卵を行う。受精卵は雄によって全て泡巣に収められ、稚魚が泳ぎだすまで雄により保護される。
- 産卵床(さんらんしょう)
- 産卵する場所。サケ、マス類は砂礫底を掘り、スズメダイは岩の表面の藻類や付着動物を歯で削り取って作る。
- 産卵記号(さんらんきごう、英: spawning mark)
- 鱗上に刻まれる繁殖期の痕跡。すべての魚類にみられるわけではない。産卵期になると成熟に伴う代謝量の増加、摂食活動の低下(停止)に伴い、鱗の表面や骨質層に吸収現象が生じる。産卵期が終わり、通常の生活に戻ると、吸収部の外側に新しい隆起線が形成され、吸収部の乱れた隆起線は痕跡として残る。
 チャネル諸島北部のアメリカナヌカザメ(トラザメ科)の卵。卵殻は三次卵膜である。 チャネル諸島北部のアメリカナヌカザメ(トラザメ科)の卵。卵殻は三次卵膜である。
 クマノミ類の卵。 クマノミ類の卵。
- 浮性卵(ふせいらん、英: pelagic egg)
- 卵の比重が周りの水の比重よりも小さく、水中を浮遊する卵。海水魚がほとんど。うち表層を浮遊する浮性卵を表層浮性卵という。
- 凝集浮性卵(ぎょうしゅうふせいらん)
- 表層浮性卵の一つ。寒天物質に包まれ、塊となって浮遊する卵。アンコウ類、ミノカサゴ類など。
- 分離浮性卵(ぶんりふせいらん、英: free pelagic egg)
- 表層浮性卵の一つ。産み出されたのち、1個ずつ分離して浮遊する卵。マアナゴ・マアジ・マイワシ・マダイ(以上、油球がある)、カタクチイワシ・マダラ・イシガレイ(以上、油球がない)など。
- 中深層浮性卵(ちゅうしんそうふせいらん)
- 外洋の中心層を浮遊する卵。ニギス、タチウオなどのもの。
- 沈性卵(ちんせいらん、英: demersal egg)
- 卵の比重が周りの水の比重よりも大きく、水底に沈んだり、他物に付着する卵。浮性卵より相対的に大きい。淡水魚、沿岸魚にみられる。
- 粘着沈性卵(ねんちゃくちんせいらん)
- 粘着卵の一つ。卵表面に粘着性の物質または付着器があり、他物に付着でき、浮力のない卵を言う。コイ・フナ類(粘液で付着)、アユ・シラウオ・ワカサギ・ハゼ・ギンポ類(付着器を備える)などの卵。
- 粘着塊状卵(ねんちゃくかいじょうらん)
- 粘着卵の一つ。産み出された卵の表面に粘着糸があり、自然にまたは親魚によって卵塊が形成される。粘液で付着する塊状卵にハタハタ、アイナメ、クジメのものなどがある。
- 纏絡卵(纒絡卵、てんらくらん)
- 付属糸で海藻などに絡まる卵。メダカ、サンマ、サヨリ、トビウオなどの卵。
- 粘着系(ねんちゃくけい、英: adhesive filament)
- 卵の表面の糸状物。他物に粘着させる役割を持つ。
- 油球(ゆきゅう、英: oil drop)
- 卵黄の中にある球形の脂肪の塊。卵の比重を軽くする。
生活・行動
- 特化(とっか)
- 原始的なものから進化するが、特に特異な特徴を進化させた場合に用いる。
- 個体変異(こたいへんい、英: indivisual variation)
- 個体間にみられる形質の違い。遺伝的なものとそうでないものがある。
- 体色変異(たいしょくへんい)
- 同種であっても体色が生息場所などによって著しく変化していること。
- 年齢(ねんれい)
- 通常、産卵期を境とする満年齢で数えるが、数え年で数える場合もある。耳石、鱗、主鰓蓋骨などに現れる年輪で数える。
- 天然魚(てんねんぎょ、英: native fish) または 在来魚(ざいらいぎょ)
- もとからその場所に生息していた居着きの魚。人間の手を借りず自然繁殖を続けているもの。
- 放流魚(ほうりゅうぎょ、英: stocker)
- 生まれが孵化場、養魚場の人工種苗魚。発眼卵、稚魚、幼魚が、あるいは未成魚、成魚に育てられてから放流された魚。
- ハッチェリーフィッシュ(英: hatchery fish)
- 特に、稚魚放流された放流魚。稚魚を放流したものが成長し回帰してきたものもこれに当る。「ハッチェリー」(英: hatchery)は孵化場の意。
- (移植による)野生繁殖魚(やせいはんしょくぎょ、英: wild fish, wild propagator, planted fish, ailen) または 自然繁殖魚(しぜんはんしょくぎょ)
- 放流された魚が自然繁殖したもの。最初の魚は放流魚なので移植群だが、生態として考えるならその子孫は野生魚である。移植魚が在来魚と異なる種の場合、それが生態を乱すという意見もある。野生繁殖魚の中には天然魚との交雑も含まれている。
- 肺呼吸(はいこきゅう、英: pulmonary respiration)
- 肺を用いてガス交換する外呼吸。呼吸運動により肺内に入ってきた空気と血管内の血液との間でガス交換がおこなわれる。魚類ではハイギョ類や一部の硬骨魚類に見られる。脊椎動物の肺は原始的な硬骨魚類の咽頭が膨出することによって生じたものである。硬骨魚類の鰾はそれが二次的に転じたとされる。
- 鰓呼吸(えらこきゅう、英: branchial respiration)
- 鰓を用いて水中でガス交換する外呼吸。水(呼吸媒質)は鰓の表面を流れる。魚類では呼吸水の流れの方向は二次鰓弁の毛細血管を流れる血液の方向と反対である。よって二次鰓弁を流れる血液と水との間には常に一定の酸素分圧の勾配が維持され、血液は水によって常に効果的な酸素飽和の仕事がなされる。
- 鰓呼吸型循環系(えらこきゅうがたじゅんかんけい、英: gill-plan of circulatory system)
- 鰓呼吸する脊椎動物にみられる血管系。円口類、魚類、両生類の幼生が行う。
- 食性(しょくせい、英: food habit, diet)
- 何を食べるかということ。胃内容物を見れば食べているものがわかる。同じ種でも成長の程度、餌の量、生息場所、時間帯、季節などによって変化する。食性によって口も変化する。肉食性、魚食性、藻食性、雑食性などという。
- 雑食魚(ざっしょくぎょ、英: omnivorous fish)
- 植物性、動物性両方の餌を食べる(雑食性)魚類。ただし、肉食魚でも草食魚でもともに異なる食餌をとることがあり(つまり肉食魚は植物餌、草食魚は動物餌)、厳密には区別は困難。典型的な雑食魚はオイカワ、コイ、キンブナ、ドジョウ、メジナ、スズメダイ類、ミゾレチョウチョウウオ、アイゴなど。
- 魚食魚(ぎょしょくぎょ、英: piscivorous fish)
- もっぱら魚を餌とする魚のこと。肉食魚の一つ。
- 濾過摂食(ろかせっしょく、英: filter feeding)
- 水中に懸濁している食物の粒子を繊毛や剛毛の動きによって集めて食べる摂食法のこと。ニシンやサバなどのプランクトン食の魚類は鰓耙を篩のようにして小甲殻類などを濾しとって食べる。表層の海水でさえ1L当り0.2-1.8mgの有機物粒子しか含まないので、濾過摂食を行う動物は大量の水を摂食器官で通過させる必要がある。
 アカククリ(マンジュウダイ科)の幼魚。有毒のヒラムシ類に擬態している。 アカククリ(マンジュウダイ科)の幼魚。有毒のヒラムシ類に擬態している。
 海藻に擬態しているリーフィーシードラゴン。 海藻に擬態しているリーフィーシードラゴン。
- 擬態(ぎたい、英: mimicry/mimesis)
- 動物が体やその一部の色彩や形態を他の物に似せること。例えば、流れ藻に暮らすツバメウオの幼魚は枯葉や藻が漂うように擬態している(隠蔽的擬態)。またアカククリの幼魚は毒性のあるヒラムシ類に擬態している(標識的擬態)。
- 隠蔽的擬態(いんぺいてきぎたい、英: mimesis) または 模倣(もほう、英: mimesis)
- 動物の形態、色彩や行動が捕食者の関心をひかない他の動物や無生物に似ているような擬態。これを隠蔽(いんぺい)やカムフラージュ(カモフラージュ、英: camouflage)に含めることもあるが、捕食者がそれを背景から識別できないようなものを隠蔽、識別しているものの食べられないものと認知するような場合を隠蔽的擬態と区別することもある。隠蔽的動物擬態(英: zoomimesis)、隠蔽的植物擬態(英: phytomimesis)、隠蔽的異物擬態(英: allomimesis)などがあり、リーフィーシードラゴン、ウィーディーシードラゴンなどは海藻と紛らわしい扁平な長い突起を持つため隠蔽的植物擬態の一つである。
- 標識的擬態(ひょうしきてきぎたい、英: mimicry)
- 動物が警告色を持つほかの動物など、捕食者の注意を引くものに似るような擬態。狭義の擬態。似せようとする側を擬態者(ぎたいしゃ、英: mimic)、似せる対象をモデル(英: model)という。美味で捕食されやすい被食者が有毒もしくは不味で警告色を持つ他の動物に姿を似せて敵を欺くベイツ擬態(ベイツぎたい、英: Batesian mimicry)と、2種以上の動物が持つ警告色がよく似た斑紋や色彩に収斂し未経験者の捕食者に食われる率を低め合うミュラー擬態(ミュラーぎたい、英: Müllerian mimicry)がある。
- 種内擬態(しゅないぎたい、英: intraspecific mimicry)
- 標識的擬態のうち、モデルが同種の他個体である擬態。口内哺育をする魚類の中には、雄の腹鰭に自種の卵そっくりな模様(エッグスポット)をもって、それを波うたせることにより雌の産卵を促す。この場合モデルは自種の卵で、信号受信者である雌を騙している。雌擬態も種内擬態の一つである。
- 雌擬態(めすぎたい、英: female mimicry)
- 種内擬態の一つ。同種の雌に擬態すること。縄張りを持てない雄が雌と似た姿を持つことで他の雄の縄張りに潜み、繁殖に加わろうとする。
- 隠蔽色(いんぺいしょく、英: concealing coloration, cryptic coloration)
- 動物個体の存在を背景の中に埋没させてしまう効果を持つ体色。捕食者は獲物に警戒されず近づけ、被食者は捕食者の眼を逃れやすくなる(保護色)。隠蔽的擬態もこれに含まれる。魚類の中にはカレイ類など周囲の色の変化に際して急速な体色変化を起こし、相応する隠蔽色を現すものもある。
- 保護色(ほごしょく、英: protectove coloration, protecting color)
- 被食者が持つ隠蔽色。捕食者の眼を逃れやすくする効果がある。寄生虫などには全く効果を持たない。捕食者が持つ隠蔽色も含める場合がある。
- カウンターシェイディング(カウンターシェーディング、英: counter shading)
- 隠蔽色の一種で、中層を遊泳する魚類の腹側が白っぽく背側が暗い色をしていること。
- 威嚇行動(いかくこうどう、英: threat behavior)
- 敵を嚇して追い払う行動。魚類では鰓蓋を広げるなどして、体を大きく見せると同時に歯や角のような武器を露わにする。通常は実際の闘争に至ることなく、その目的を達する。
- 儀式的闘争(ぎしきてきとうそう、英: conventional fighting) または 試合闘争(しあいとうそう、独: Turnierkampf)
- 同種個体間における闘争で、一定のルールに従って行っているように見えるもの。多くは最後には弱い方が服従行動をとって相手のそれ以上の攻撃を抑制するか逃げ去るかして一方または双方が傷ついたり死ぬことなく終わる。攻撃は儀式的闘争のために発達したと考えられる器官によって相手の体の特定の部位(トゲウオでは強固な皮膚を持つ顎)だけに与えられ、致命傷を与える部位には向けられない。スポーツの試合に似た性格を持つ。ゲーム理論に基づいて個体の利益を見る観点から説明されている。
- 縄張り(なわばり、英: territory)
- 他の個体または集団の生息や侵入を許さない一定の生息区域。
- 順位(じゅんい、英: order)
- 社会集団の中の個体の優位序列。魚類ではベラ類に多くみられる。ハーレムの雄がいなくなると最高順位の雌が雄に性転換する。その他ハナダイ、ヤッコの仲間でも知られ、クマノミ類では逆の例がみられる。クマノミは単独の大型の雌がイソギンチャクを中心に一定の縄張りを持ち、複数の雄が随うが、産卵に参加できるのは順位が最高の雄のみである。雌がいなくなると、最上位の雄が雌に性転換する。
- ハレム(はれむ、ハーレム、英: Harem)
- 大きなオスのなわばりの中に数尾のメスがすむ一夫多妻のグループ。魚においては、性転換する種の一部でみられ、ベラ科やキンチャクダイ科などでみられる。最大個体のオスが死ぬと、ハレムで次に大きいメスがオスに性転換する[45]。
- 発音魚(はつおんぎょ、英: sound-producting fish)
- 遊泳、採餌、呼吸などに伴い生じる音は除き、警戒、求愛、威嚇、認識などの目的で音を発する魚の総称。発音魚の多くは鰾が大きい役割を果たす。その他、歯、肩帯、鰭条なども発音に利用される。
- 発音(はつおん)
- 発音の目的は敵への威嚇や仲間の位置の察知、同種同士の警戒などがあるが、ドジョウ類の呼吸音など生態的意味を持たないものも多い。カサゴでは縄張りの侵入者を威嚇するときに、ニベ類では産卵期に激しく発音する。
- 摩擦音(まさつおん)
- 体の一部を摩擦する音。ゴンズイの場合、胸鰭の棘を付着している骨に押し付ける。ヒイラギは主上顎骨の上端から後方に伸びている突起を顎骨と擦り合せる。アジ類は上下咽頭骨を、マツカサウオやイトヨは腹鰭の大きな棘を立てその基部の関節面に摩擦する。セミホウボウは頭骨中の舌顎骨を他の頭骨と摩擦し、カジカは鰓蓋の一部を摩擦し、フグ目の多くは上下の歯を擦りあわせて発音する。ドンコなどでも発音のための特別な筋肉は発達していないが、肩帯の骨の振動音などを鰾で増幅させて音を出す。
- 振動音(しんどうおん)
- 鰾に付随する筋肉の収縮で鰾が振動して非常に高い音を発する。ホウボウ、イシダイ、シマイサキ、コトヒキ、ニベ類、カサゴ類などが振動音を発音する。イットウダイ、ハタンポ、ニベ類、シマイサキ、カサゴなどでは鰾と頭骨、脊椎骨、体腔壁を結ぶ発音筋が発達し、それにより鰾壁を振動させて大きな音を出す。マトウダイ、ガマアンコウ、カナガシラなどでは鰾壁に密着した発音筋が発達し、その働きにより発音する。特にホウボウの雄は咽頭骨を摩擦し、鰾を共鳴器とするため声が高い。
- 呼吸音(こきゅうおん)
- ドジョウやタウナギなど腸呼吸を行う魚が発する。水中に酸素が少なくなったり、空気中に出したりすると空気を口から消化管の中に入れ、腸壁に分布する毛細血管を吸入し、残物を肛門から出す。これが狭い腸管を漏れる際音を発する。
- 棲み分け(すみわけ、英: habitat segregation, interactive habitat segregation)
- 似た生活様式を持つ2種以上が同じところに棲むこともできるのに生息場所を分け合っている現象。例えばイワナとヤマメは夏季の水温13℃の付近を境にして分かれて棲むことが多い。しかし他種のいない場合はヤマメは13℃より水温の低い上流にも、イワナは13℃より水温の高い下流にも生息する。
- 片利共生(へんりきょうせい、英: commensalism)
- 共生者の一方が何らかの利益を受け、もう一方は利益も不利益も受けていない状況のこと。
- 掃除魚(そうじうお、英: Cleaner fish)
- クリーニング行動を行う魚類の総称。例としてベラ科やコバンザメ科などがある。
- クリーニング行動(くりーにんぐこうどう、英: Cleaning symbiosis)または 掃除行動(そうじこうどう)、クリーニング(くりーにんぐ)
- 生物の相利共生における行動の1つで、死んだ皮膚組織や外部寄生虫を採食することをいう。クリーニング行動を行う魚類を掃除魚と呼ぶ。他種の体表や口の中をつつき、外部寄生虫や、傷んだ皮膚、体表粘液などを食べる。幼魚のみこの行動を行う種も多い[45]。
- クリーニングステーション(くりーにんぐすてーしょん、英: Cleaning station)
- クリーニング行動はそれを行う掃除魚のなわばり内の決まった場所で行われ、クリーニングをしてほしい魚はそこを訪れることから、そのような場所を特にこう呼ぶ[45]。
- ラム換水(ラムかんすい、英: ram ventilation)
- 魚類の呼吸法の一つ。能動的な呼吸運動をせず、自身の移動の結果として口から入ってくる水流により鰓に一方向性の水流を与えて呼吸する方法。サバ、サメ類にみられる。
- 比重調節(ひじゅうちょうせつ、英: regulation of specific gravity)
- 魚類は遊泳力を使うか自身の比重を変えることにより自由に生息深度を変えられる。サメ、エイ、カレイやヒラメなど鰾を持たない魚類は遊泳をやめるとその深度を維持できず、その比重により浮き上がるか沈むかしてしまう。
- 発光魚(はっこうぎょ、英: luminous fish)
- 発光魚はツノザメ目、フウセンウナギ目(フクロウナギ科など)、ソトイワシ目(ソコギス亜目など)、ワニトカゲギス目(ムネエソ科、ヨコエソ科などほぼすべて)、キュウリウオ目(セキトリイワシ科など)、ハダカイワシ目(ハダカイワシ科、ソトオリイワシ科など)、タラ目(ソコダラ科など)、アンコウ目(チョウチンアンコウ科など)、キンメダイ目(マツカサウオ科など)、スズキ目(ヒイラギ科、テンジクダイ科など)などの分類群に現れる。
- 発光(はっこう、英: luminescence) または 生物発光(せいぶつはっこう、英: bioluminescence)
- 発光する魚はすべて海産で光の色は淡緑色や白色のものが多い。中には濃緑色や深紅色(キュウリエソなど)のものもある。発光の目的は認識信号(仲間に自分の所在を知らせる)、照明(自分の進路を照らす)、誘引(餌生物を誘き寄せる)、威嚇(敵を嚇す)、防御(敵の眼を眩ます)など種類によってさまざま。
- 自力発光(じりきはっこう) または 閉鎖式(へいさしき)
- 自分で発光素を分泌し、酸化による発光。細胞から発光素ルシフェリンを分泌しそれが体内もしくは水中の酸素で燃焼して起こる。このとき酵素ルシフェラーゼにより燃焼が補助される。この燃焼は炎も熱も発生しない冷光である。
- ほとんどが深海魚だが、ツマグロイシモチの腸管の一部は自力発光腺となっている。深海魚ではハダカイワシ科、ヨコエソ科、ムネエソ科、カラスザメ属(カラスザメやフジクジラ)などである。発光器は眼のような構造をし、体に散在している。しかしカラスザメ類は皮膚器官として無数にあり、ミズテングでは発光器がないのにも拘らず発光する。ハダカイワシ類のものはよく発達しており、神経や血管もきわめて多く発達しており、瞬間的に発光できる。チョウチンアンコウ類では擬餌状体に発光器がある。
- 共生発光(共棲発光、きょうせいはっこう、英: symbiotic luminescense) または 他力発光(たりきはっこう)、開孔式(かいこうしき)
- 発光器の中に発光バクテリアが共生し、その発光によって光を発する。発光バクテリアが侵入するために発光腺は必ず消化管のある部分か肛門付近に開いている。ヒイラギ(食道を取り巻く部分が発光腺)、マツカサウオ(下顎の前端に左右一対の発光器))、ホタルジャコ(魚体の下半部全体が発光器、発光腺は肛門付近に開孔)、トウジン科の多く(腹部に縦走した細長い発光腺、後端は肛門直線に開孔)などがこの仕組みを持つ。
 発電魚のデンキウナギ(デンキウナギ目ギュムノートゥス科)。 発電魚のデンキウナギ(デンキウナギ目ギュムノートゥス科)。
 弱発電魚のギュムノートゥス属の一種 Gymnotus inaequilabiatus(デンキウナギ目ギュムノートゥス科) 弱発電魚のギュムノートゥス属の一種 Gymnotus inaequilabiatus(デンキウナギ目ギュムノートゥス科)
- 発電魚(はつでんぎょ、英: electric fish) または 電気魚(でんきうお)
- 発電器官を備えた魚類。系統的には少なくとも6回独立に出現しており[46]、進化における収斂の例として知られる。発電力の強さにより2つに大別できる。発電魚は発電器官を使って短い時間間隔で自らの周囲に電場を形成し、その電場の乱れを感知して餌や外敵の存在を知る。
- 強い発電力を持つ発電魚
- シビレエイ Narke japonica、シビレエイの一種 Torpedo marmorata、デンキナマズ Malapterurus electricus、デンキウナギ Electrophorus electricus、ミシマオコゼの仲間 Astroscopus など。数百Vにも及ぶ。その起電力は、他の動物を倒すという攻撃的、または防御的役割を果たす。
- 弱発電魚(じゃくはつでんぎょ) または 弱電気魚(じゃくでんきうお)
- モルミルス Mormyrus やギュムノートゥス Gymnotus、ガンギエイ Raja など。1V程度の放電を50–750 Hz程度の頻度で出す。これらの魚では別に電気を受容する感覚器官(電気受容器)を備えており、放電によって周囲に形成される電場を感知し(電場定位)、外敵や障害物の接近を知る方向探知機構として用いられる。
- 魚介毒(ぎょかいどく、英: marine toxin)
- 生きている魚介類が持っている自然毒の総称。
 刺咬毒をもつミナミゴンズイ(ゴンズイ科)。最悪の場合死に至る。 刺咬毒をもつミナミゴンズイ(ゴンズイ科)。最悪の場合死に至る。
- 刺咬毒(しこうどく)
- 一般に高分子蛋白質だが、充分な毒液が集めにくく、毒の成分が不安定だったりすることなどにより性状については明らかでない点が多い。ゴンズイやカサゴ類が持つ。毒腺から分泌され、腫脹、痛み、痙攣、呼吸困難などを起こし、致命的な場合もある。
- 刺毒魚(しどくぎょ、英: venomous fish)
- 体表の一部の棘に毒腺を持ち、他の生物が刺されると毒液が注入される。約200種程度が知られ、ゴンズイやオコゼ類では刺さると先端が折れ毒が外的に注入されやすくなっている。
- 皮膚毒(ひふどく)
- 多くはペプチドを含む分子量数千の物質。ヌノサラシ、アゴハタ、コバンハゼ、ミナミウシノシタ、ハコフグなどが持つ。表皮の毒分泌細胞やこの細胞が真皮に落ち込んでできた毒腺に貯蔵される。他個体を殺し、防御的効果を持つ。
- フグ毒(ふぐどく、河豚毒、鰒毒、英: fugu toxin)
- フグの臓器(卵巣と肝臓に顕著)に含まれる毒素。食中毒の一種。細菌の産生する毒素をフグが蓄えたもので、食用フグに多く含まれるが、カナフグやキタマクラなどではほとんど認められない。他に皮膚や腸から毒が認められるものもあるが、血液はほぼ無毒である。フグ以外にもツムギハゼやカリフォルニアイモリの卵、タコからも認められる。
- 毒成分はテトロドトキシン(英: tetrodotoxin、TTX)で、毒作用は細胞膜における神経や骨格筋の電位依存性ナトリウムチャネルのナトリウムイオン透過経路の阻害による。症状は筋肉弛緩、感覚麻痺、嘔吐、神経節遮断(血圧下降・腸管の運動抑制)で呼吸麻痺による中毒死を起こす。類似の毒素にチリキトキシン(ヤセヤドクガエルから)サキシトキシン(貝毒)などがある。
- シガテラ毒(シガテラどく、英: ciguatera toxin)
- 熱帯・亜熱帯地域に生息する魚類を食べて起こる食中毒の総称。その魚に固有の毒ではなく、有毒プランクトンで産生され、食物連鎖を通じて蓄積されたもの。主な毒性を持つ物質としてシガトキシン(英: ciguatoxin、CTX)とアルテリンなどが分離されている。バラフエダイ、バラハタ、ドクカマス、ドクウツボなどの内臓や筋肉に存在する。致死率は低く、手や口の周りの感覚異常や運動失調などの神経系障害を引き起こす。その他下痢や嘔吐、ドライアイスセンセーション(英: dry-ice sensation)、関節痛、倦怠感などを引き起こす。
- イクシオトキシン(英: ichthyotoxin)
- 魚雷型(ぎょらいがた)
- 魚の遊泳方法の基本形は3つに分けられ、その遊泳法の一つ。紡錘形の魚の遊泳。体の大部分はほとんど屈伸せず、尾鰭と体の後端部を左右に屈伸する。体側筋が発達している。
- 漕艇型(そうていがた)
- 遊泳法の一つ。対鰭(特に胸鰭)によって水を後方に押して前進する。魚雷型に比べると遅く、持続力も弱い。エイ類にみられる。
- この型の変形で、背鰭と臀鰭のみを波状に動かすか左右に振って推進するものがある。フグ目にみられる。
- 蛇行型(だこうがた)
- 遊泳法の一つ。延長形の魚にみられる。長い体を左右に何度も屈伸して水を押し前進する。尾鰭の発達は悪く、持たないものもある。対鰭は遊泳に与らないため対鰭を持たないものもある。ヒラメ類は体が扁平のためヒルのように主軸を垂直に屈伸して前進する。
- 回遊(回游、洄游、かいゆう、英: migration)
- 魚が地理的に大きく移動すること。餌を求めたり、産卵場所を求め移動したり、生息に適した場所を求めて移動したりと、一つの場所にとどまらず移動を繰り返す。
- 通し回遊(とおしかいゆう) または (広義の)両側回遊(りょうそくかいゆう)
- 海と川を往来する回遊。
- 遡河回遊(そかかいゆう)
- 長期間海で成長した後、産卵のため川を遡ること。遡河回遊魚も参照。
- 降海回遊(こうかいかいゆう) または 降河回遊(こうかかいゆう)
- 長期間川で成長し、産卵のため海へ下ること。降河回遊魚も参照。
- 両側回遊(りょうそくかいゆう)
- ある一定期間海でも川でも生活し、産卵に関係なく領域を移動すること。普通河川で孵化し、流下して仔稚魚期を海で過ごし、稚魚期中後期ごろ遡上し、成長して河川で産卵するものが多い。両側回遊魚も参照。
- 河川回遊(かせんかいゆう)
- 河川の中での回遊。孵化した仔魚が河川中を流下し、遡上しながら成長すること。ソウギョ、オイカワなど。
- 海洋回遊(かいようかいゆう、英: oceanic migration)
- 海洋中で行われる回遊。沿岸回遊、外洋表層回遊、垂直回遊などが含まれる。
- 偶来回遊(ぐうらいかいゆう)
- 偶発的に海から河川に遡上すること。スズキ、ボラ、ギンガメアジ、ナガエバなどがこれに当る。
- 索餌回遊(さくじかいゆう)
- 餌を食べるための回遊。
- 索餌回遊域(さくじかいゆういき)
- 餌を求めて回遊(索餌回遊)する海域。
- 陸封型(りくふうがた、英: land locked form) または 陸封魚(りくふうぎょ)
- 元来は河川や湖沼と海を行き来していた魚が温度などの気候や地形変化などで淡水域に閉じ込められ河川や湖で一生を送るようになったもの。サケ科、トゲウオ科にみられる。
- 降海型(こうかいがた、英: sea run form) または 走海型(そうかいがた)、シーラン(英: sea run form)
- 生活の中心が淡水域にある種において、海に下って生活しているグループ。ウグイなどにみられる。
 ベニザケ(サケ科)の遡上。 ベニザケ(サケ科)の遡上。
- 遡上(そじょう)
- 海と川を行き来する魚が海から川へ遡ること。湖に棲む魚が川へ遡る際にも使われる。
- 食物連鎖において周囲の生態系と一体化して栄養をつなぎ、重要な役割を果たしている。
- モドリ(戻り、もどり)
- 海と川を行き来する魚において、海へ降りられる環境でも流程が長い河川の場合、途中で引き返してくることがあり、その個体。
- 母川回帰(ぼせんかいき) または サーモン・ラン(英: salmon run)
- サケ科魚類が一度外洋に出て生まれ育った川に回帰(英: homing)すること。太陽コンパス、磁気コンパス(英: magnetic compass)、嗅覚を組み合わせて利用していることが分かっている。うち嗅覚が最も重要である。
生息水域による魚の種類
 淡水魚のミナミメダカ(メダカ科)。 淡水魚のミナミメダカ(メダカ科)。
- 淡水魚(たんすいぎょ、英: freshwater fish)
- 淡水に棲む魚。魚類全体の41.8%。
- 純淡水魚(じゅんたんすいぎょ) または (広義の)一次淡水魚(いちじたんすいぎょ)
- 一生淡水域に限り生息している魚。
- 一次性淡水魚(いちじせいたんすいぎょ) または 一次淡水魚(いちじたんすいぎょ)
- 古い地史年代から淡水域で種の分化を生じ、現在では海水にまったく耐性のないもの。ハイギョ、コイ科、ナマズ目魚類の大部分やチャラシン科など。魚類全体の33.1%(約10450種)。
- 二次性淡水魚(にじせいたんすいぎょ) または 二次淡水魚(にじたんすいぎょ)
- 淡水域に限り生息しているが、海水に対する耐性が多少あり、短期間なら海水中を移動できるもの。メダカ科、カワスズメ科魚類など。--魚類全体の8.7%。
- 通し回遊魚(とおしかいゆうぎょ、英: diadromous fish) または (広義の)両側回遊魚(りょうそくかいゆうぎょ)
- 通し回遊(両側回遊)する魚。本来は海水中で分化してきた分類群に属するが、生活史のある期間を海と川の両方に持ち、両者間を往来できるもの。
- 遡河回遊魚(そかかいゆうぎょ、英: anadromous fish)
- 淡水で生まれ、海で育った後、産卵のために淡水に戻る魚。サケ、マス類の多くはこれに当る。陸封がみられる。
- 降河回遊魚(こうかかいゆうぎょ、英: catadromous fish)
- 産卵のために海へ下り、海に生まれた仔稚魚は淡水域に入り、そこで育つ魚類のこと。ウナギなど。
- 両側回遊魚(りょうそくかいゆうぎょ、英: amphidromous fish)
- 淡水に生まれすぐに海に下り、産卵とは関係なく再び淡水に戻る魚。アユ、チチブ、ヨシノボリ、ボウズハゼ、アユカケなど。汽水域や河口域を往復する魚類も含める考え方もある。陸封がみられる。アユなどを淡水性両側回遊魚、ボラなどを海水性両側回遊魚という。
- 海水魚(かいすいぎょ)
- 海水に棲む魚。魚類全体の58.2%(14550種)。
- 汽水魚(きすいぎょ)
- 内湾から沿岸の、塩素量1–17%の範囲に生活の中心を持つ魚。サッパ、コノシロ、ドロクイ、サヨリ、ボラ、マハゼ、シマハゼ、ウロハゼなど。
- 沿岸魚(えんがんぎょ、英: coastal fish)
- タイドプール域から水深50 mまでの沖合に分布する魚。外洋魚の対語。沿岸付近ではチョウチョウウオ科、ベラ科、ブダイ科、イシダイ科、キス科など。やや沖合ではニシン科、エソ科、サバ科、アジ科、ヒラメ科など。
- 近海魚(きんかいぎょ) または 沿海魚(えんかいぎょ)
- 水深50mから大陸棚縁辺または大陸斜面上部にわたる範囲に生息する魚。沿岸魚との明確な区別は困難で沿岸魚に含まれることも多い。表層魚は沿岸魚とよく似た組成で、底層はマダイ、ヘダイ、コチ類、ヒメコダイ、カスミサクラダイ、ニギス、アオメエソ類、アラなど。
- 外洋魚(がいようぎょ)
- 大陸棚上にも侵入してくるが、生活の本拠地を外洋に持つ魚。表層性魚類はシイラ、カツオ、マグロ類、カジキ類、大型サメ類など。および、中深層、深層性の深海魚。
- 表層遊泳魚(ひょうそうゆうえいぎょ) または 表層魚(ひょうそうぎょ)
- 上部表層から下部表層に棲む魚。カツオ・マグロ類など。
- 中深層遊泳魚(ちゅうしんそうゆうえいぎょ) または 中深層魚(ちゅうしんそうぎょ)
- 中深層に生息する魚。ヨコエソ科、ハダカイワシ科など。
- 深層遊泳魚(しんそうゆうえいぎょ) または 深層魚(しんそうぎょ)
- 上部漸深層以下に生息する魚。ミツマタヤリウオ、チョウチンアンコウなど。
 深海底に生息している底生魚、シンカイエソ Bathysaurus mollis(シンカイエソ科)。水深4,900 mでの観察例がある。非常に高い水圧がかかっている。 深海底に生息している底生魚、シンカイエソ Bathysaurus mollis(シンカイエソ科)。水深4,900 mでの観察例がある。非常に高い水圧がかかっている。
- 底生魚(ていせいぎょ) または 底魚(そこうお)
- 底帯に棲む魚。底生生物(ベントス、英: benthos)のうちのメガベントス(巨大ベントス、英: megabenthos)に分類される。また底生動物(動物ベントス、英: benthic animal)[注釈 3]のうち遊泳性底生動物(英: nektobenthos)[注釈 4]に分類される。
- 底層遊泳魚(ていそうゆうえいぎょ)
- 底帯を遊泳する魚。
- 回遊魚(かいゆうぎょ、英: migratory fish) または 回遊性魚類(かいゆうせいぎょるい)
- マグロ、カツオ、ニシン、サンマなど、餌や産卵場所を求めて広く移動する魚。
- 鹹水魚(かんすいぎょ)
- 浮き魚(浮魚、うきうお、英: pelagic fish)
- 底魚の対語。表層や中層に棲む魚の総称。浅海から深海に及ぶ広大な水域に分布し、分類群も多様。底魚との明確な境界はない。深度帯によって表層性、中深層性、漸深層性、深海性などに大別される。浮魚は食物連鎖の上位に位置し、水平もしくは鉛直的回遊を行うものが多いが、特に表層性のものにおいて顕著。つまり発育段階や生態的周期性との関連で生活様式や地理的分布の性状が著しく動的な構造を持つ場合が多い。イワシ、サケ、マグロなどの多獲性の水産的に重要な魚種はここに含まれる。中深層性のものは小型で、種多様性の高いハダカイワシやヨコエソ類などマイクロネクトン魚類が中心である。これらは海洋における鉛直的物質輸送の役割が注目され、直接漁獲対象となることはまれである。
- 深海魚(しんかいぎょ、英: deep-sea fish)
- 深海に生息する魚。
- 冷水種(れいすいしゅ、英: cold-water species)
- 生態分布において、冷水中だけにみられる種。生活の最低温度、最適温度、最高温度が低温にあり、高緯度の海域や湖沼、高山の湖沼に多く、河川では上流域にみられる。海水魚ではタラやニシン、淡水魚ではイワナやマス類など。
地形と海洋生態系
 ポルトガル、Póvoa de Varzimのタイドプール。干潮時でも海水を残っており、稚魚や磯魚の格好の生育場となる。 ポルトガル、Póvoa de Varzimのタイドプール。干潮時でも海水を残っており、稚魚や磯魚の格好の生育場となる。
- タイドプール(英: tide pool) または 潮溜り(潮だまり、しおだまり)
- 主に潮間帯で干潮時でも海水が残っているところ。潮間帯特有の小形の魚や、沖に棲む魚の幼魚や稚魚がみられる。
- 外洋(がいよう、英: open sea) または 外洋域(がいよういき、英: oceanic region, oceanic zone)
- 水深200 m以深の沖合一帯。岸から沖合に向かっての水平区分のうちの一つ。陸からの栄養が届かず、栄養分的に貧しい。複雑な地形が少なく、単純な環境なため魚種が少なく、一つの種が大きな群れを作り遊泳していることが多い。遊泳力の優れた魚がみられる。
- 沖合(おきあい、英: off-shore)
- 陸方に対し、外海部のことを言う。
- 沿岸(えんがん、英: coast) または 沿岸域(えんがんいき、英: littoral zone)
- 水深200 m以浅の岸部。岸から沖合に向かっての水平区分のうちの一つ。海浜域(かいひんいき)と沿岸域に二分されるがどちらも潮汐、塩分濃度、海藻の発達など環境多様性を反映し、魚類相は豊富。熱帯域ではサンゴ礁性魚類が、温帯域ではカレイ類、タラ類などが代表種。サケ・マス類は産卵接岸時に沿岸魚となる。
- 大陸棚(たいりくだな、英: continental shelf) または 陸棚(りくだな)
- 水深200 m以浅の海底。
- サンゴ礁(珊瑚礁、さんごしょう、英: coral reef)
- 炭酸カルシウムの骨格を大量に生産する造礁サンゴ、有孔虫、石灰藻などの造礁生物(ぞうしょうせいぶつ、英: hermatypic prganism)の骨格が集積してできたサンゴ礁石灰岩が海面近くまで達して防波構造物となる地形。サンゴ礁は造礁サンゴを中心に魚や甲殻類の棲み家となり、海洋生態系の中で最も種多様性の高いサンゴ礁群集(珊瑚礁群集、さんごしょうぐんしゅう、英: coral reef vommunity)が形成される。環礁も参照。
- 礁湖(しょうこ、英: lagoon, coral lake)
- サンゴ礁に取り囲まれた水域。礁湖が形成されるサンゴ礁は主に、陸地とサンゴ礁の外縁が数km離れる堡礁(ほしょう、英: barrier reef)である。
- 岩礁(がんしょう、英: rock reef)
- 根(ね)
- 平坦な砂泥などの海底から立ち上がった岩。周辺は魚の格好の棲み家となる。回遊魚でも沖の海底にある根にやってきて、そこを餌場とするものもいる。根の周囲に棲む魚を根付きの魚という。
- 砂底(さてい)
- 海底の底質の用語。砂の海底。磯と比較すると単純で、生息している生物の種数は少ない。ソウハチ、アオミシマ、クサフグなど砂に身を隠す生物が多く見られる。
- 砂泥底(さでいてい)
- 砂交じりの泥からなる海底。沿岸から離れた岩礁は平坦な砂泥底となる。ハタハタやキス、コチ、ホウボウ、ヒラメ・カレイ類など多くの底生魚が生息する。
- 砂礫底(されきてい)
- 小石交じりの砂からなる海底。
- 遮蔽海岸(しゃへいかいがん)
- 湾奥などで外洋水の交流が少ない海岸。
 河川とその河口(天竜川) 河川とその河口(天竜川)
- 河川(かせん、英: river) または 川(河、かわ)
- 淡水のうち流水域。河川に棲む生物の群集を河川の群集(かせんのぐんしゅう、英: stream community, lotic community)といい、河川に生息している生物の生活様式、生理的特性、種類組成や環境条件を総合して識別された河川の区分を河川の生態的区分(かせんのせいたいてきくぶん、英: ecological division of the river)という。日本ではアマゴ域、オイカワ域、コイ域の区分がある。同様の区分として、欧米諸国では古くからトラウト域(英: trout region)、グレイリング域(英: grayling region)、バーベル域(英: barbel region)、ブリーム域(英: bream region)に区別されていた。
- 瀬(せ、英: rapid)
- 河川のうち流水性が卓越し底質が岩石からなる部分。
- 淵(渕、ふち、英: riverpool, stream pool)
- 河川のうち止水性が強く砂泥の堆積する部分。
- 湖沼(こしょう、英: lake) または 湖(みずうみ、英: lake)、池(いけ、英: pond)、沼(ぬま、英: marsh)
- 湖と沼の区別は厳密でなく、規模により分けることが多い。規模が大きい場合湖流(こりゅう)が発生し、河川と似た環境となる。夏季には高水温となることが多い。代表的な生息魚種はコイ、ギンブナ、オイカワ、アカヒレタビラ、タナゴ、モツゴ、ナマズ、メダカなどである。
- 感潮域(かんちょういき)
- 川の中で、潮の干満の影響を受ける区域。汽水域より広い意味でつかわれる。
- 汽水域(きすいいき)
- 河川から流入する淡水と海水が混じっている水域。内湾に流れ込む大きな河川の河口では、広い範囲にわたって汽水域ができ、川が運んできた有機物に富んだ土砂が堆積し干潟が形成される。ここには魚の餌となる底生生物が多く、これを食物として一生汽水域で過ごす魚や、沖に棲む魚の仔稚魚などが多く見られる。
- 砕波帯(さいはたい)
- 波打ち際と砕ける波の沖側限界との間を言う。アユなど多くの種の仔稚魚期の生育場として重要である。
- 藻場(もば、英: seaweed bed)
- 大型の水生植物や藻類が群落を作っている場所。アマモが生育する藻場をアマモ場、ホンダワラの生育する藻場をガラモ場という。
 サルガッソ海を漂う流れ藻。 サルガッソ海を漂う流れ藻。
- 流れ藻(ながれも、英: drifting seaweed, floating seaweed)
- 褐藻類やホンダワラなどの大型の海藻が岩場から離れたりや本体からちぎれて海中に出て浮遊したもの。産卵場や幼魚の育成場となる。生息地としての意味を持ち、一つの生態系が構成されている。ツバメウオなどは流れ藻に産卵する。サルガッソ海ではホンダワラが浮漂したまま繁殖を続けており、そこに生活するのに適応した形態の魚類や甲殻類が分化している。
- マングローブ(英: mangrove)
- 海山(かいざん、英: seamount)
- アルカリ栄養湖(アルカリえいようこ、英: alkalitrophic lake)
- カルシウム含有量の特に多い湖。アフリカ中央地溝帯など大陸内部の乾燥地域にみられ、アルカリ性は炭酸ナトリウムの解離による。生産力は大きいが非調和的。生物の種類は少なく、アルスロスピラ属 Arthrospira など藍藻類が水の華(英: water-bloom)を形成し、動物プランクトンはワムシ類が生息する。魚類はティラピアおよびヒレナマズ属 Clarias が生息している。
- 人工魚礁(じんこうぎょしょう、英: artificial reef, artificial fishing reef)
- 集魚の目的で人工的に構築した海中構造物。天然の魚礁を模して海底に沈設することもおおいが、最近は浮体を係留する浮魚礁(うきぎょしょう、パヤオ、英: payaos)も増えてきた。船を沈めるなどして集魚することは江戸時代から行われており、築磯などと呼ばれてきた。1952年から国の補助事業として実施され、1954年から魚類を対象する人工魚礁とそれ以外の築磯に分けられたのがこの用語が公式に使われた最初である。魚礁に魚が集まるのは餌、隠れ場のほかに渦流効果や陰影効果のためといわれている。
脚注
注釈
- ^ なお、両生類と爬虫類も5対、鳥類・哺乳類では4対。有羊膜類では鰓は形成されず、咽頭囊は閉じる。
- ^ 網膜上の錐体細胞や桿体細胞の規則的配列のこと。
- ^ ベントスのうち動物のものを指す。
- ^ 遊泳力を持ち遊泳と底生の相互移行的な生活をする。
出典
参考文献
- 阿部宗明『原色魚類大圖鑑』(初版)北隆館、1987年11月25日。ISBN 4832600087。
- 岡村収、尼岡邦夫『山溪カラー名鑑 日本の海水魚』(3版第8刷)山と溪谷社、2009年3月10日、14–31, 32, 34頁。ISBN 9784635090278。
- 巌佐庸、倉谷滋、斎藤成也、塚谷裕一『第5版 岩波生物学辞典』(第5版)岩波書店、2013年2月26日、38–1515頁。ISBN 9784000803144。
- 能勢幸雄『魚の事典』(初版)東京堂出版、1989年1月30日、27–430頁。ISBN 4490102453。
- 蒲原稔治、岡村収『原色日本海水魚図鑑I』(初版)保育社、1985年7月31日、xiii–xvi頁, 76–147頁。ISBN 4586300728。
- 桜井淳史、渡辺昌和『淡水魚ガイドブック』永岡書店、2008年11月10日、4, 240–243頁。ISBN 9784522213544。
- 井田齊、奥山文弥『サケマス・イワナのわかる本』(初版)山と溪谷社、2012年7月5日、42–63, 202–226頁。ISBN 9784635360746。
- 中坊徹次『日本産魚類検索 全種の同定 第三版 I』(第3版)東海大学出版会、2013年、19, 21, 22, 25頁。ISBN 9784486018049。
- 木村義志『フィールドベスト図鑑7 日本の海水魚』GAKKEN、2000年8月4日、4–5, 15, 250–252頁。ISBN 4054011217。
- 小西英人『釣魚1400種図鑑』(初版)エンターブレイン、2011年3月28日、5–7頁。ISBN 9784047271807。
- 仲谷一宏『SHARKS サメ ―海の王者たち―』(初版)ブックマン社、2011年9月1日、9, 102, 123–127, 143–145頁。ISBN 9784893087539。
- 小林道信『熱帯魚・水草 カラー図鑑』西東社、2004年3月15日、196-198頁。ISBN 9784791606689。
- 黒木真理、塚本勝巳『旅するウナギ 1億年の時空をこえて』(初版)東海大学出版会、2011年7月20日、40–41, 83, 103頁。ISBN 9784486019077。
- 著者:ビバマンボ・北村雄一、監修:三宅裕志・佐藤孝子『DVD-ROM&図解 動く!深海生物図鑑』講談社 ブルーバックス、2010年8月20日、148–149頁。ISBN 9784062576918。
- 林原自然科学博物館『OCEAN! 海はモンスターでいっぱい(特別展カタログ)』読売新聞大阪本社企画事業部、2011年、9–21頁。
- 中島淳『LOACHES OF JAPAN 日本のドジョウ 形態・生態・文化と図鑑』山と渓谷社、2017年、27頁。
関連項目
外部リンク
|
|







_diagram_cropped.GIF)






.png)











,_Parsley_Bay,_Sydney.jpg)

















.jpg)




_mouthbrooding.jpg)



.JPG)





-1.jpg)



