DNA topoisomerases (or topoisomerases) are enzymes that catalyze changes in the topological state of DNA, interconverting relaxed and supercoiled forms, linked (catenated) and unlinked species, and knotted and unknotted DNA.[1][2] Topological issues in DNA arise due to the intertwined nature of its double-helical structure, which, for example, can lead to overwinding of the DNA duplex during DNA replication and transcription. If left unchanged, this torsion would eventually stop the DNA or RNA polymerases involved in these processes from continuing along the DNA helix. A second topological challenge results from the linking or tangling of DNA during replication. Left unresolved, links between replicated DNA will impede cell division. The DNA topoisomerases prevent and correct these types of topological problems. They do this by binding to DNA and cutting the sugar-phosphate backbone of either one (type I topoisomerases) or both (type II topoisomerases) of the DNA strands. This transient break allows the DNA to be untangled or unwound, and, at the end of these processes, the DNA backbone is resealed. Since the overall chemical composition and connectivity of the DNA do not change, the DNA substrate and product are chemical isomers, differing only in their topology.
Discovery
The first DNA topoisomerase was discovered in bacteria by James C. Wang in 1971 and was initially named ω (omega) protein;[3] it is now called Escherichia coli (E. coli) topoisomerase I (topo I) and is a representative of the type IA family of enzymes. Subsequently, a similar activity was found in eukaryotic cells (rat liver) by James Champoux and Renato Dulbecco;[4] the enzyme responsible, eukaryotic topo I, has a distinct mechanism and is representative of the type IB family. The first type II topoisomerase to be discovered was DNA gyrase from bacteria, by Martin Gellert and coworkers in 1976,[5] and also characterized by Nicholas Cozzarelli and co-workers.[6] DNA gyrase catalyzes the introduction of negative supercoils into DNA and is the only type II enzyme to do this, all the others catalyze DNA relaxation. Type II enzymes are mechanistically distinct from type I in being ATP-dependent and transiently cleaving both DNA strands rather than just one. Type II topoisomerases were subsequently identified from bacterial viruses and eukaryotes.[7][8][9] Topo EC-codes are as follows: ATP-independent (type I), EC 5.6.2.1; ATP-dependent (type II): EC 5.6.2.2. The exception among the type I topoisomerases, reverse gyrase, which contains a helicase domain (EC 3.6.4.12) and introduces positive supercoiling in an ATP-dependent manner. Therefore it is the sole type I topoisomerase classified as EC 5.6.2.2 (Table 1).
DNA topology
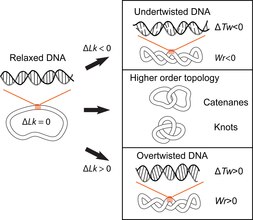
The double-helical structure of DNA involves the intertwining of the two polynucleotide strands around each other, which potentially gives rise to topological problems. DNA topology refers to the crossing of the two DNA strands that alters the twist of the double helix and gives rise to tertiary conformations of DNA, such as supercoils, knots and catenanes.[10] Potential topological issues associated with the double-helical structure of DNA were recognized soon after its structure was first elucidated in 1953 by James Watson, Francis Crick and Rosalind Franklin[11][12][13] and developed further by the work of Max Delbruck and John Cairns.[14][15] Closed-circular double-stranded DNA can be described by 3 parameters: Linking number (Lk), Twist (Tw) and Writhe (Wr) (Fig. 1). Where Lk refers to the number of times the two strands are linked, Tw refers to the number of helical turns in the DNA, measured relative to the helical axis, and Wr quantifies the coiling of the path of the DNA helix in space and is often equated with 'supercoiling'.
Figure 1. Overview of DNA topology. The linking number (Lk) describes the number of times that the two single-strands of DNA cross each other in a closed circular DNA molecule. Relaxed DNA molecules have an intrinsic linking number (Lk0) corresponding to the twisting of the two single-strands around each other in the double helix, approximately once per 10.5 base-pairs (Tw0~ 10.5). Supercoiling corresponds to increases or decreases of the linking number (∆Lk) that result from cellular processes such as DNA transcription and replication (Figure 2). Changes in linking number are accommodated by changes in twist (torsion) (Tw) and writhe (Wr), which change the structure and mechanics of DNA. Knotting within a DNA molecule and links between DNA molecules (catenanes) represent higher order topological conformations of DNA.
The 3 parameters are related as follows: Lk = Tw +Wr. This is a mathematical identity originally obtained by Călugăreanu in 1959[16] and is referred to as the Călugăreanu, or Călugăreanu–White–Fuller, theorem.[17][18] Lk cannot be altered without breaking one or both strands of the helix; Tw and Wr are interconvertible and depend upon the solution conditions. Supercoiling is a vernacular term for DNA with a non-zero linking difference, more correctly referred to as specific linking difference (σ = ΔLk/Lk0, where Lk0 is the mean linking number of the relaxed DNA circle). DNA is said to be positively supercoiled if Lk of it is higher than Lk0 for the relaxed state (Lk-Lko = ΔLk, ΔLk>0); that means that Tw and/or Wr are increased relative to the relaxed molecule. Conversely, DNA is negatively supercoiled if Lk of the molecule is lower than the Lk0 (ΔLk<0).
The consequences of topological perturbations in DNA are exemplified by DNA replication during which the strands of the duplex are separated; this separation leads to the formation of positive supercoils (DNA overwinding or overtwisting) ahead of the replication fork and intertwining of the daughter strands (precatenanes) behind[10][19] (Fig. 2). If the positive supercoils are not relaxed, progression of the replication fork is impeded, whereas failure to unlink the daughter strands prevents genome segregation, which is required for cell division.[20] Transcription by RNA polymerase also generates positive supercoiling ahead of, and negative supercoiling behind, the transcriptional complex (Fig. 2). This effect is known as the twin-supercoiled domain model, as described by Leroy Liu and James Wang in 1987.[21] These topological perturbations must be resolved for DNA metabolism to proceed, allowing the cell to efficiently replicate, transcribe and partition the genome to enable cellular division and vitality. Knots in DNA can be found in bacteriophages and as products of recombination reactions.[10] In general, knots in DNA are detrimental and need to be removed (by topoisomerases). DNA catenanes are formed upon the replication of circular molecules and need to be resolved by topoisomerases or recombinases to allow proper separation of daughter molecules during cell division. In addition to the detrimental aspects of DNA topology that require resolution, there are also beneficial aspects. For example, plasmid replication requires negative supercoiling of the origin, which facilitates local melting and exposes single-stranded DNA required to initiate replication. Similarly, initiation of replication from the main bacterial origin oriC also requires negative supercoiling.[22][23] Furthermore, compaction of the E. coli genome is achieved in part by negative supercoiling.
Figure 2. Topological consequences of DNA metabolism. i) During DNA replication, strand separation leads to positive supercoiling ahead of the advancing protein machinery, and precatenane formation behind. Precatenanes form as the newly-synthesised duplexes wrap around one and other, and, if not removed prior to complete of replication, catenated DNA molecules are formed. ii) During transcription, strand separation leads to positive supercoiling ahead of the advancing protein machinery, and negative supercoil formation behind.[1]
Types
DNA topoisomerases are enzymes that have evolved to resolve topological problems in DNA (Table 2).[10] They do this via transient breakage of one or both strands of DNA. This has led to the classification of topos into two types: type I, which catalyze reactions involving transient single-stranded breaks, and type II, which catalyze reactions involving transient double-stranded breaks (Fig. 3; Table 2). Sub-types exist within these classifications.
Figure 3. Summary of topoisomerase types and catalytic mechanisms. The topoisomerases are categorized based on whether they catalyze single- (type I) or double-stranded (type II) DNA breaks. The type I topoisomerases are further subdivided to type IA, IB and IC. Type IA form a transient covalent bond to the 5ʹ DNA phosphate and function via a strand passage mechanism. Type IB and IC form a transient covalent bond to the 3ʹ DNA phosphate and function via a controlled-rotation mechanism. Type II topoisomerases are further subdivided into type IIA and IIB. Both form a transient covalent bond to the 5ʹ DNA phosphate of both strands of the duplex and function via a strand-passage mechanism.
Type I
These enzymes catalyze changes in DNA topology via transient single-stranded breaks in DNA. Reactions can occur on both single- and double-stranded DNA substrates and can proceed via a 'swivel' or 'strand-passage' mechanism (Fig. 3). The range of reactions includes: DNA supercoil relaxation, unknotting of single-stranded circles, and decatenation, provided at least one partner has a single-stranded region. In the case of the archaeal enzyme, reverse gyrase, positive supercoiling of DNA is possible.[24]
Type IA
Type IA are monomeric and bind to single-stranded segments of DNA. They introduce a transient single-stranded break through the formation of a tyrosyl-phosphate bond between a tyrosine in the enzyme and a 5′-phosphate in the DNA. The segment of DNA within which the break occurs is called the 'gate' or G-segment, and its cleavage allows the passage of another segment of DNA, the 'transport' or T-segment, to be passed through in a 'strand-passage' process.[25] This is followed by ligation of the G-segment. For strand passage to occur, topo IA must undergo a conformational change to open the DNA gate and allow T-segment transfer. During a DNA relaxation reaction this process changes the linking number of the DNA by +/-1 (Fig. 4). Examples of type IA topoisomerases include prokaryotic topo I and III, eukaryotic topo IIIα and IIIβ and the archaeal enzyme reverse gyrase. Reverse gyrase, which occurs in thermophilic archaea, comprises a type IA topo coupled to a helicase, and is the only known enzyme that can introduce positive supercoils into DNA.[24] The gene encoding reverse gyrase is also found in some groups of thermophilic bacteria, where it was likely transferred by horizontal gene transfer from Archaea.[26]
Figure 4. Strand-passage mechanism for type IA topos. (1) Topo binds the G-segment ssDNA region, (2) the G-segment is cleaved. (3) The topo DNA gate is opened, (4) which allows T-segment transfer through the cleaved G-strand. (5) The DNA gate is closed, (6) and the G-strand is re-ligated, changing the linking number by 1. (7) The topo can then go through another round of relaxation or dissociate from the DNA. Type IA topo (domains 1-4) is in pink, the active site tyrosine is yellow and the DNA is grey.
Type IB
Type IB topoisomerases catalyze reactions involving transient single-stranded breaks in DNA through the formation of a tyrosyl-phosphate bond between a tyrosine in the enzyme and a 3′-phosphate in the DNA. Rather than utilizing a strand-passage mechanism, these enzymes operate via a 'swivel' or 'controlled rotation' of the cleaved strand around the intact strand.[27] This controlled-rotation mechanism was first described for Vaccinia topo I[27][28] and permits DNA rotation of the free end around the intact strand, the speed being controlled by 'friction' within the enzyme cavity, before the nick is re-ligated (Fig. 3). This results in a variable change of linking number per cleavage and religation event. This mechanism is distinct from that of the type IA enzymes, and the two groups of enzymes are structurally and evolutionarily unrelated. Examples of type IB topoisomerases include eukaryotic nuclear and mitochondrial topo I in addition to viral topo I, though they have been identified in all three domains of life.
Type IC
Type IC topoisomerases share a similar mechanism to the type IB enzymes but are structurally distinct. The sole representative is topo V, found in the hyperthermophile Methanopyrus kandleri.[29]
Type II
Type II topoisomerases catalyze changes in DNA topology via transient double-stranded breaks in DNA. Reactions occur on double-stranded DNA substrates and proceed via a strand-passage mechanism (Fig. 5). The range of reactions include DNA relaxation, DNA supercoiling, unknotting, and decatenation. Whereas all type II topoisomerases can catalyze DNA relaxation, gyrase, an archetypal bacterial topoisomerase, can also introduce negative supercoils. In contrast to type I topoisomerases that are generally monomeric, type II topoisomerases are homodimers or heterotetramers. They are classified into two subtypes based on evolutionary, structural, and mechanistic considerations. The general strand-passage mechanism for the type II topos begins with the binding of one DNA duplex, termed the gate segment (G-segment), at the DNA gate. Another duplex, termed the transport segment (T-segment), is captured by an ATP-operated clamp and passed through a transient break in the G-segment, involving 5ʹ phosphotyrosine linkages in both strands, before it is released through the C-gate and the G-segment is re-ligated (Fig. 5). Enzyme turnover requires the binding and hydrolysis of ATP.
Figure 5. Type II topoisomerase strand-passage mechanism. (1) G-segment is bound at the DNA-gate and the T-segment is captured. (2) ATP binding stimulates dimerization of the N-gate, the G-segment is cleaved and the T-segment is passed through the break. (3) The G-segment is re-ligated and the T-segment exits through the C-gate. For type IIB topos, there is no C-gate so once the T-segment passes through the G-segment, it is released from the enzyme. (4) Dissociation of ADP and Pi allows N-gate opening, a scenario where the enzyme either remains bound to the G-segment, ready to capture a successive T-segment, or (5) dissociates from the G-segment.
Type IIA
Type IIA topoisomerases catalyze transient double-stranded breaks in DNA through the formation of tyrosyl-phosphate bonds between tyrosines in the enzyme (one on each subunit) and 5′-phosphates staggered by 4 bases in opposite DNA strands. The strand-passage reaction can be intra- or intermolecular (Fig. 5), thus permitting changes in supercoiling and knotting, or unlinking, respectively. This process changes the linking number of the DNA by +/-2. Examples of type IIA topoisomerases include eukaryotic topo IIα and topo IIβ, in addition to bacterial gyrase and topo IV. DNA gyrase conforms to the same double-strand passage mechanism as other type II enzymes but has unique features connected with its ability to introduce negative supercoils into DNA. The G segment is part of a much longer piece of DNA (>100 bp) that is wrapped around the enzyme, one arm of which forms the T-segment that is passed through the double-stranded break (Fig. 5). In the case of gyrase, a substantial amount of the free energy from ATP hydrolysis is transduced into torsional stress in DNA, i.e. supercoiling is an energy-requiring process.[30] Further, in the absence of ATP, gyrase is able to remove negative supercoils in a slower DNA relaxation reaction.
Type IIB
Type IIB also catalyze transient double-stranded breaks through the formation of tyrosyl-phosphate bonds between tyrosines in the enzyme and 5′-phosphates in opposite strands of the DNA, but in the case of IIB enzymes the double-stranded breaks have a 2-base stagger. Type IIB enzymes show important structural differences, but are evolutionarily related to the type IIA enzymes. These differences include the lack of one of the protein 'gates' (the C gate) (Fig. 5). Originally found in archaea, they have also been found in eukaryotes, and, in particular, in plants; examples include topo VI and topo VIII. Topo VI is the best-studied enzyme of this sub-type and is thought to be a preferential decatenase.[31]
As drug targets
For the non-specialist perhaps the most important aspect of topoisomerases is their role as drug targets both for antibacterial and anti-cancer chemotherapy; several topoisomerase-targeted antibacterial and anti-cancer drugs are listed among the 2019 World Health Organization Model List of essential Medicines. The reason for this prominence is that their reactions proceed via transient breaks in DNA, which, if stabilized by drug binding, can lead to cell death due to the generation of toxic single- or double-stranded breaks in genomic DNA. The majority of topo-targeted drugs act in this way, i.e. they stabilize the enzyme-DNA covalent cleavage intermediate.[32][33][34]
Antibacterial compounds
Although type I topos, such as bacterial topo I, are viable antibiotic targets,[35] there are currently no compounds in clinical use that target these enzymes. However, the type II enzymes, DNA gyrase and DNA topoisomerase IV, have enjoyed enormous success as targets for the widely-used fluoroquinolone antibiotics, (Fig. 6).
Fluoroquinolones (FQs)
Quinolone antibacterial compounds were first developed in the 1960s and have been in clinical use since the 1980s.[36] FQ derivatives, such as ciprofloxacin, levofloxacin and moxifloxacin (Fig. 6) have been highly-successful. These compounds work by interacting with their target (gyrase or topo IV) and DNA at the cleavage site to stabilize the DNA-protein covalent cleavage intermediate. Specifically, they intercalate into the DNA and prevent the DNA religation step of the topoisomerase reaction (Fig. 5). This is a highly-effective mechanism of inhibition that is also used by several topoisomerase-targeted anti-cancer drugs. Despite their spectacular success, resistance to FQs is a serious problem.[36] A variety of other compounds, such as quinazolinediones and imidazolpyrazinones,[37] work in a similar manner and it is hoped that some of these will replace FQs in the future.
Aminocoumarins
Aminocoumarins (Fig. 6), such as novobiocin, clorobiocin and coumermycin A1, are natural products from Streptomyces that inhibit the ATPase reaction of gyrase and topo IV.[37] Although they can be very potent against their target, they suffer from permeability and toxicity issues, and thus have not enjoyed the level of clinical success of the FQs.
Proteinaceous inhibitors
There are a number of protein inhibitors of gyrase, including the bacterial toxins CcdB, MccB17, and ParE,[38][39][40] that stabilize the cleavage complex, in a similar manner to FQs. Although these proteins are not viable as antibacterials, their mode of action could inspire the development of novel antibacterial compounds. Other protein inhibitors of gyrase prevent DNA binding by the topoisomerase rather than stabilizing cleavage complexes. These include YacG[41] and pentapeptide repeat proteins, such as QnrB1 and MfpA;[42][43] these protein inhibitors also confer resistance to fluoroquinolones.
Figure 6. Structures of antibiotic compounds that target bacterial DNA gyrase and topoisomerase IV.
Anti-cancer compounds
Both human topo I and topo II (both α and β isoforms) can be targeted in anticancer chemotherapy (Fig. 7).[32][33][44][45][46][47] Most of these compounds act in a similar way to FQs, i.e. by stabilizing the DNA-protein covalent cleavage complex; for this they have become known as topoisomerase poisons, distinct from catalytic inhibitors.[33][34][48] Several human topoisomerase inhibitors are included on the World Health Organization's List of Essential Medicines.
Camptothecin (CPT)
Camptothecin (Fig. 7), originally derived from the tree Camptotheca acuminata, targets human topo I and derivatives such as topotecan and irinotecan are widely used in cancer chemotherapy.[33] Camptothecin and its derivatives act by stabilizing the topo I cleavage complex, preventing religation of the protein-mediated nick in the DNA. These interfacial inhibitors are stabilized by stacking interactions with the nicked DNA and hydrogen bonding to the enzyme. Although CPT derivatives stabilize a single-strand cleavage complex, subsequent collisions with replication or transcription machinery are thought to generate toxic double-stranded DNA breaks. These compounds are used as first or second line therapies to treat cancers including colorectal, ovarian, lung, breast, and cervical. However, CPT derivatives suffer from limitations associated with toxicity and limited therapeutic half-lives due to chemical instability. New topo I inhibitors, the indenoisoquinolines and fluoroindenoisoquinolines, overcome the limitations of CPT derivatives and are currently in clinical trials.[49]
Etoposide (VP-16)
Etoposide (Fig. 7) and its close relative teniposide (VM-26) are epipodophyllotoxin derivatives obtained from the rhizome of wild mandrake that target topo II by stabilizing the covalent cleavage complex and preventing religation of the cleaved DNA.[46] These are typically used in conjunction with other chemotherapy drugs to treat cancers including testicular tumors, small-cell lung cancer, and leukemia. Etoposide treatment can result in secondary leukemias arising from specific genomic translocations, mainly involving topo IIβ.[46]
Doxorubicin
Figure 7. Structures of antitumor compounds that target human topoisomerases.
Doxorubicin (Fig. 7) and the related derivatives daunorubicin, epirubicin, and idarubicin are anthracyclines obtained from the bacterium Streptomyces[48] that target human topo II, stabilizing the cleavage complex in a similar manner to other topoisomerase poisons. Mitoxantrone is a synthetic anthracenedione that is chemically and functionally similar to anthracyclines.[47] The anthracyclines were the first topoisomerase inhibitors used to treat cancer and remain among the most widely employed and effective treatments for a broad range of cancers including breast cancer, lymphoma, leukemias, carcinomas, sarcomas, and other tumors.[47] These compounds are DNA intercalating agents and as such can impact a wide range of cellular DNA processes in addition to specifically poisoning topo II.[33] Additional cytotoxicity stems from redox reactions involving anthracyclines that generate reactive oxygen species. Generation of reactive oxygen, along with poisoning of topo IIβ, result in the dose-limiting cardiotoxicity of the anthracyclines.[33]
Merbarone
Merbarone is a thiobarbituric acid derivative, and dexrazoxane (ICRF-187), one of several related bisdioxopiperazine derivatives, (Fig. 7) are examples of catalytic inhibitors of topo II, i.e. they prevent completion of the catalytic cycle of topo II but do not stabilize the DNA cleavage complex. Whereas these catalytic inhibitors exhibit cytotoxicity and have been tested in clinical trials, they are not currently in clinical use for cancer therapy.[47] However, dexrazoxane, which blocks ATP hydrolysis by topo II, is used to prevent cardiotoxicity associated with the anthracyclines.[50][51]
Topoisomerase
Subfamily type
Function
Multimericity
Metal dependence
ATP dependence
Single- or double-stranded cleavage?
Cleavage polarity
Change in link number (L)
Topoisomerase I
(E. coli)
Type IA
Removes (-), but not (+) supercoils. Prevents excessive supercoiling of the genome, and supports transcription
Monomer
Yes (Mg2+)
No
SS
Topoisomerase III
(E. coli)
Removes (-), but not (+) supercoils; overlapping function with topoisomerase IV
Removes (-), but not (+) supercoils; assists in the unlinking of precatenanes in cellular DNA replication; can catalyze the knotting, unknotting, and interlinking of single-stranded circles as well as the knotting, unknotting, catenation, and decatenation of gapped or nicked duplex DNA circles
Role in suppressing recombination or supporting transcription in neurons
Homodimer
Topoisomerase VI
(Archaea)
Type IIB
Relaxes (+) and (-) supercoils; responsible for decatenating replication intermediates
Heterotetramer
Yes (Mg2+)
Yes
DS
5'
±2
Role of topoisomerase in transcriptional regulation
At least one topoisomerase, DNA topoisomerase II beta (topo IIβ), has a regulatory role in gene transcription. Topo IIβ–dependent double-strand DNA breaks and components of the DNA damage repair machinery are important for rapid expression of immediate early genes, as well as for signal-responsive gene regulation.[52][53][54][55] Topo IIβ, with other associated enzymes,[54] appears to be important for the release of paused RNA polymerase at highly transcribed or long genes.[56][57][58]
Topo IIβ in initiation of transcription
Stimulus-induced DNA double-strand breaks (DSBs) that are limited to a short-term (10 minutes to 2 hours) are induced by topo IIβ in the promoter regions of signal-regulated genes. These DSBs allow rapid up-regulation of expression of such signal responsive genes in a number of systems (see Table below). These signal-regulated genes include genes activated in response to stimulation with estrogen, serum, insulin, glucocorticoids (such as dexamethasone) and activation of neurons. When the induced DNA double-strand break has been repaired, then transcription of the signal-responsive gene returns to a low basal level.[52]
Paused RNA polymerase and limited, short-term topo IIβ-induced DNA double-strand break. The 5' ends of DNA are covalently joined to tyrosine within the topo IIβ dimer-PARP-1 complex. The non-homologous end joining DNA repair pathway components DNA-PKcs, Ku70/Ku80 and DNA ligase are also closely associated with the topo IIβ dimer-PARP-1 complex.
Topo IIβ and PARP-1 were found to be constitutively present at a moderate level near the transcription start site of a promoter of a signal-responsive gene. After the signal occurred, topo IIβ caused a double-strand break and PARP-1 was involved in replacing histone H1 by HMGB1/HMGA2, which can promote transcription.[55] Topo IIβ and PARP-1 increased at the site of the double-strand break and components of the non-homologous end joining DNA repair pathway, including DNA-PKcs, Ku70/Ku80 and DNA ligase IV assembled with topo IIβ and PARP-1. This assemblage was all present at the linker DNA adjacent to a single nucleosome in the promoter region of a gene (see Figure). The nucleosome was close to the transcription start site of the gene.[55] The components of the non-homologous end joining DNA repair pathway were essential to the closing of the DNA double-strand break.[52]
Agents causing temporary DNA double-strand breaks (DSBs) required for transcription and proteins associated with the DSBs in signal-responsive genes.
RNA polymerase II frequently has a pausing site that is about 30–60 nucleotides downstream of
the transcription start site of a gene.[64][65] The pausing of RNA polymerase II at these sites and the controlled release of the pausing is thought to have a regulatory role in gene transcription. As pointed out by Singh et al.,[58] "about 80% of highly expressed genes in HeLa cells are paused". Very short-term, but not immediately resealed, topo IIβ-induced DNA double-strand breaks occur at sites of RNA polymerase II pausing, and appear to be required for efficient release of the paused state and progression to gene transcription.[56][57][58] For the genes at which it occurs, the DNA double-stranded break induced by TOP2B is thought to be part of the process of regulation of gene expression.
^Wang JC (February 1971). "Interaction between DNA and an Escherichia coli protein omega". Journal of Molecular Biology. 55 (3): 523–533. doi:10.1016/0022-2836(71)90334-2. PMID4927945.
^Baldi MI, Benedetti P, Mattoccia E, Tocchini-Valentini GP (June 1980). "In vitro catenation and decatenation of DNA and a novel eucaryotic ATP-dependent topoisomerase". Cell. 20 (2): 461–467. doi:10.1016/0092-8674(80)90632-7. PMID6248247. S2CID42645648.
^Cairns J (March 1963). "The bacterial chromosome and its manner of replication as seen by autoradiography". Journal of Molecular Biology. 6 (3). Elsevier: 208–213. doi:10.1016/s0022-2836(63)80070-4. PMID14017761.
^White JH (1969). "Self-Linking and the Gauss Integral in Higher Dimensions". American Journal of Mathematics. 91 (3): 693–728. doi:10.2307/2373348. ISSN0002-9327. JSTOR2373348.
^Sundin O, Varshavsky A (September 1981). "Arrest of segregation leads to accumulation of highly intertwined catenated dimers: dissection of the final stages of SV40 DNA replication". Cell. 25 (3): 659–669. doi:10.1016/0092-8674(81)90173-2. PMID6269752. S2CID24408315.
^Liu LF, Liu CC, Alberts BM (March 1980). "Type II DNA topoisomerases: enzymes that can unknot a topologically knotted DNA molecule via a reversible double-strand break". Cell. 19 (3): 697–707. doi:10.1016/s0092-8674(80)80046-8. PMID6244895. S2CID8921868.
^ abMaxwell A, Bush NG, Germe T, McKie SJ (2018). "Non-quinolone topoisomerase inhibitors". In Fong IW, Drlica K (eds.). Antimicrobial resistance and implications for the twenty-first century. New York: Springer. pp. 593–618. ISBN978-0-387-72417-1. OCLC227210110.
^Pommier Y, Tanizawa A, Kohn KW (1994). "Mechanisms of topoisomerase I inhibition by anticancer drugs". DNA Topoisomerases: Topoisomerase-Targeting Drugs. Advances in Pharmacology. Vol. 29B. Elsevier. pp. 73–92. doi:10.1016/s1054-3589(08)61132-1. ISBN978-0-12-032930-4. PMID8996602.
^ abcdMurphy MB, Mercer SL, Deweese JE (January 2017). "Inhibitors and Poisons of Mammalian Type II Topoisomerases". Advances in Molecular Toxicology. Vol. 11. Elsevier. pp. 203–240. doi:10.1016/b978-0-12-812522-9.00005-1. ISBN9780128125229.
^ abHande KR (October 1998). "Clinical applications of anticancer drugs targeted to topoisomerase II". Biochimica et Biophysica Acta (BBA) - Gene Structure and Expression. 1400 (1–3): 173–184. doi:10.1016/s0167-4781(98)00134-1. PMID9748560.
^Speyer JL, Green MD, Kramer E, Rey M, Sanger J, Ward C, et al. (September 1988). "Protective effect of the bispiperazinedione ICRF-187 against doxorubicin-induced cardiac toxicity in women with advanced breast cancer". The New England Journal of Medicine. 319 (12): 745–752. doi:10.1056/nejm198809223191203. PMID3137469.
^Cvetković RS, Scott LJ (2005). "Dexrazoxane: a review of its use for cardioprotection during anthracycline chemotherapy". Drugs. 65 (7): 1005–1024. doi:10.2165/00003495-200565070-00008. PMID15892593.
Wang JC (2009). Untangling the Double Helix: DNA entanglement and the action of the DNA topoisomerases. Cold Spring Harbor: Cold Spring Harbor Laboratory Press. p. 245. ISBN978-0-87969-879-9.
У Вікіпедії є статті про інших людей із прізвищем Химинець. Химинець Василь ВасильовичХиминець Василь Васильович Народився 13 листопада 1970(1970-11-13) (53 роки)с. Кибляри Ужгородський район, Закарпатська область, Українська РСР, СРСРГромадянство СРСР→ УкраїнаНаціональн...
Ermont Koordinat: 48°59′32″N 2°15′37″E / 48.9922°N 2.2603°E / 48.9922; 2.2603NegaraPrancisArondisemenPontoiseKantonErmont (ibukota)AntarkomuneCA Val et Forêt Ermont merupakan sebuah komune di pinggiran utara Paris, Prancis. Terletak 17.2 km (10.7 mil) dari pusat kota Paris. Demografi Menurut sensus 1999, populasinya 27.494. Perkiraan untuk 2005 adalah 28.300 jiwa. Angkutan Ermont dilayani oleh stasiun Ermont – Eaubonne yang merupakan persimpangan pa...
Mayor of JacksonvilleSeal of the City of JacksonvilleFlag of the City of JacksonvilleIncumbentDonna Deegansince July 1, 2023StyleThe HonorableTerm length4 years, renewable once consecutivelyInaugural holderWilliam J. MillsFormation1832Salary$206,218Websitewww.coj.net/mayor/meet-the-mayor Elections in Florida Federal government Presidential elections 1848 1852 1856 1860 1868 1872 1876 1880 1884 1888 1892 1896 1900 1904 1908 1912 1916 1920 1924 1928 1932 1936 1940 1944 1948 1952 1956 1960...
Kawasan AnalanjirofoKawasanLetak di MadagaskarNegara MadagaskarLuas • Total21.930 km2 (8,470 sq mi)Populasi (2013) • Total1.035.132 • Kepadatan47/km2 (120/sq mi)Zona waktuUTC3 (EAT) Analanjirofo adalah sebuah kawasan di timur laut Madagaskar. Sampai 2009, kawasan tersebut merupakan bagian dari Provinsi Toamasina. Kawasan tersebut berbatasan dengan Kawasan Sava di utara, Kawasan Sofia di barat, Kawasan Alaotra-Mangoro di barat d...
This article does not cite any sources. Please help improve this article by adding citations to reliable sources. Unsourced material may be challenged and removed.Find sources: Kampung Sungai Kayu Ara – news · newspapers · books · scholar · JSTOR (November 2015) (Learn how and when to remove this template message) Kampung Sungai Kayu Ara is a small village in the Petaling Jaya Utara (PJU6) section of Petaling Jaya, Selangor, Malaysia. This village is l...
Anti-Discrimination Act 1991Parliament of Queensland Long title An Act to promote equality of opportunity for everyone by protecting them from unfair discrimination in certain areas of activity and from sexual harassment and certain associated objectionable conduct. Passed3 December 1991Royal assent9 December 1991Commenced30 June 1992Introduced byHon. D.M. Wells (Murrumba – Attorney-General)Status: Current legislation The Queensland Anti-Discrimination Act 1991 is an act of the Parliament ...
Vysšaja Liga 1977высшая лига 1977 Competizione Vysšaja Liga Sport Calcio Edizione 41ª Organizzatore FFSSSR Date dal 1º aprile 1977al 10 novembre 1977 Luogo Unione Sovietica Partecipanti 16 Formula Girone all'italiana Risultati Vincitore Dinamo Kiev(8º titolo) Retrocessioni Karpaty Kryl'ja Sovetov Kujbyšev Statistiche Miglior marcatore Blochin (17) Incontri disputati 240 Gol segnati 486 (2,03 per incontro) Cronologia della competizione ...
SobbleTokoh PokémonKarya seni oleh Ken SugimoriPermainanperdanaPokémon Sword dan Shield (2019)Pengisi suara Bahasa Inggris Erica Schroeder Bahasa Jepang Sayaka Senbongi InformasiSpesiesPokémonTipeAir Sobble, yang dikenal di Jepang sebagai Messon (メッソンcode: ja is deprecated ), adalah spesies Pokémon dalam waralaba media Pokémon milik Nintendo dan Game Freak. Pertama kali diperkenalkan dalam Pokémon Sword dan Shield dan menjadi salah satu dari tiga Pokémon yang tersedia untuk pem...
Cet article est une ébauche concernant un anthropologue brésilien. Vous pouvez partager vos connaissances en l’améliorant (comment ?) selon les recommandations des projets correspondants. Curt NimuendajúBiographieNaissance 17 avril 1883IénaDécès 10 décembre 1945 (à 62 ans)AmazonasSépulture Cemitério do Redemptor (d)Nationalités allemande (jusqu'en 1920)brésilienneActivité AnthropologueStèle commémorative dans un parc de sa ville natale, Iéna (Allemagne)modifier -...
Cet article est une ébauche concernant l’histoire. Vous pouvez partager vos connaissances en l’améliorant (comment ?) selon les recommandations des projets correspondants. Consultez la liste des tâches à accomplir en page de discussion. Le massacre de Poneriai (ou massacre de Ponary) est l'exécution en 1941 de 100 000 personnes, majoritairement juives, par les Einsatzgruppen, SD et SS allemands avec la collaboration de Sonderkommando lituaniens (Ypatingasis būrys et batai...
Pozzomaggiore Pottumajòre, PottumaggiòreKomuneComune di PozzomaggioreLokasi Pozzomaggiore di Provinsi SassariNegaraItaliaWilayah SardiniaProvinsiSassari (SS)Pemerintahan • Wali kotaMariano SoroLuas • Total78,77 km2 (30,41 sq mi)Ketinggian438 m (1,437 ft)Populasi (2016) • Total2,617[1]Zona waktuUTC+1 (CET) • Musim panas (DST)UTC+2 (CEST)Kode pos07018Kode area telepon079Situs webhttp://www.comune.pozzomaggio...
19th and 20th century dispute about popular and learned language varieties of Greek This article may be too long to read and navigate comfortably. When this tag was added, its readable prose size was 14,500 words. Consider splitting content into sub-articles, condensing it, or adding subheadings. Please discuss this issue on the article's talk page. (January 2018) The Greek language question (Greek: το γλωσσικό ζήτημα, to glossikó zítima) was a dispute about whether the vern...
Tobias PreisBorn1981NationalityGermanAlma materJohannes Gutenberg University of MainzKnown forIdentification of links between online behavior and real world economic eventsScientific careerFields Data Science Data mining Forecasting Complex Systems Computational Social Science Institutions Warwick Business School Alan Turing Institute Boston University University College London Swiss Federal Institute of Technology Zurich Massachusetts Institute of Technology Deutsche Bank Artemis ...
Measurement of distance from river mouth Part of Fourteen Mile Island in the Allegheny River - the island is named for its distance from the river's mouth in river miles. A river mile is a measure of distance in miles along a river from its mouth. River mile numbers begin at zero and increase further upstream. The corresponding metric unit using kilometers is the river kilometer. They are analogous to vehicle roadway mile markers, except that river miles are rarely marked on the physical rive...
Political party in Nigeria This article has multiple issues. Please help improve it or discuss these issues on the talk page. (Learn how and when to remove these template messages) This article needs additional citations for verification. Please help improve this article by adding citations to reliable sources. Unsourced material may be challenged and removed.Find sources: All Progressives Grand Alliance – news · newspapers · books · scholar · JSTOR (M...
إدينسون كافاني (بالإسبانية: Edinson Cavani) كافاني مع الأوروغواي في كأس العالم 2018 معلومات شخصية الاسم الكامل إدينسون روبيرتو كافاني غوميز[1] الميلاد 14 فبراير 1987 (العمر 37 سنة)سالتو، الأوروغواي الطول 1.88 م (6 قدم 2 بوصة)[2] مركز اللعب مهاجم الجنسية الأوروغواي معلومات �...
Ermenegol VIIConte d'UrgellIn carica1154 - 1184 PredecessoreErmengol VI il Castigliano SuccessoreErmengol VIII Nascita1120/30 MorteRequena, 11 agosto 1184 Luogo di sepolturaMonastero di Bellpuig de las Avellanas (Os de Balaguer) DinastiaCasa di Barcellona PadreErmengol VI il Castigliano MadreArsenda de Cabrera ConiugeDolce FigliErmengolMarchesa eMiraglia Ermengol, o Armengol o Ermengardo detto el de Valencia (poiché morì nei pressi di Valencia[1]) (1120/30 – Requena, 11 agos...
This article relies largely or entirely on a single source. Relevant discussion may be found on the talk page. Please help improve this article by introducing citations to additional sources.Find sources: UCL Prize Lecture in Life and Medical Sciences – news · newspapers · books · scholar · JSTOR (July 2013) The UCL Prize Lecture in Life and Medical Sciences (previously UCL Prize Lecture in Clinical Science) is a prize awarded annually by University C...
Monthly DC Comics comic book Booster GoldCover to Booster Gold vol. 2 #1. Art by Dan Jurgens.Publication informationPublisherDC ComicsScheduleMonthlyFormatOngoing seriesGenre Superhero Publication dateOctober 2007 – August 2011No. of issues47Main character(s)Booster GoldRip HunterSkeetsCreative teamCreated byDan JurgensWritten byKeith GiffenJ.M. DeMatteisDan JurgensGeoff JohnsJeff KatzArtist(s)Chris BatistaDan JurgensNorm Rapmund Booster Gold was an ongoing monthly DC Comics comic book seri...
I più importanti continua linguistici in Europa In linguistica, la mutua intelligibilità o intercomprensione è la proprietà che caratterizza un gruppo di lingue distinte quando i rispettivi parlanti possono capire con relativa facilità la lingua dell'altro, senza doverla apprendere.[1][2] L'intercomprensione reciproca può essere asimmetrica fra le lingue, nel caso in cui il parlante di una lingua capisca più e più facilmente la lingua dell'altro piuttosto che il contra...











")












")
