–ź—Ä–≥—Ė–Ĺ—Ė–Ĺ
–Ü–Ĺ—ą—Ė –Ĺ–į–∑–≤–ł
őī-–≥—É–į–Ĺ—Ė–ī–ł–Ĺ-őĪ-–į–ľ—Ė–Ĺ–ĺ–≤–į–Ľ–Ķ—Ä—Ė–į–Ĺ–ĺ–≤–į –ļ–ł—Ā–Ľ–ĺ—ā–į
–Ü–ī–Ķ–Ĺ—ā–ł—Ą—Ė–ļ–į—ā–ĺ—Ä–ł
–ź–Ī—Ä–Ķ–≤—Ė–į—ā—É—Ä–ł
–ź—Ä–≥, Arg, R –Ě–ĺ–ľ–Ķ—Ä CAS
7200-25-1 PubChem
232 –Ě–ĺ–ľ–Ķ—Ä EINECS
200-811-1 DrugBank
DB00125 KEGG
C02385 –Ě–į–∑–≤–į MeSH
Arginine ChEBI
29016 SMILES
NC(CCCNC(N)=N)C(O)=O InChI
InChI=1S/C6H14N4O2/c7-4(5(11)12)2-1-3-10-6(8)9/h4H,1-3,7H2,(H,11,12)(H4,8,9,10)/t4-/m0/s1 –Ě–ĺ–ľ–Ķ—Ä –Ď–Ķ–Ľ—Ć—ą—ā–Ķ–Ļ–Ĺ–į
1725413 –Ě–ĺ–ľ–Ķ—Ä –ď–ľ–Ķ–Ľ—Ė–Ĺ–į
83283
–í–Ľ–į—Ā—ā–ł–≤–ĺ—Ā—ā—Ė
–ú–ĺ–Ľ–Ķ–ļ—É–Ľ—Ź—Ä–Ĺ–į —Ą–ĺ—Ä–ľ—É–Ľ–į
C6 H14 N4 O2
–ú–ĺ–Ľ—Ź—Ä–Ĺ–į –ľ–į—Ā–į
174,2 –≥/–ľ–ĺ–Ľ—Ć
–ó–ĺ–≤–Ĺ—Ė—ą–Ĺ—Ė–Ļ –≤–ł–≥–Ľ—Ź–ī
–Ď—Ė–Ľ—Ė –ļ—Ä–ł—Ā—ā–į–Ľ–ł
–†–ĺ–∑—á–ł–Ĺ–Ĺ—Ė—Ā—ā—Ć (–≤–ĺ–ī–į )
87,1 –≥/–Ľ
–ö–ł—Ā–Ľ–ĺ—ā–Ĺ—Ė—Ā—ā—Ć (pK a )
2,488
–Ü–∑–ĺ–Ķ–Ľ–Ķ–ļ—ā—Ä–ł—á–Ĺ–į —ā–ĺ—á–ļ–į
10,75
–Ě–Ķ–Ī–Ķ–∑–Ņ–Ķ–ļ–ł
–ö–Ľ–į—Ā–ł—Ą—Ė–ļ–į—Ü—Ė—Ź –Ą–°
Xi
–Į–ļ—Č–ĺ –Ĺ–Ķ –∑–į–∑–Ĺ–į—á–Ķ–Ĺ–ĺ —Ė–Ĺ—ą–Ķ, –ī–į–Ĺ—Ė –Ĺ–į–≤–Ķ–ī–Ķ–Ĺ–ĺ –ī–Ľ—Ź —Ä–Ķ—á–ĺ–≤–ł–Ĺ —É —Ā—ā–į–Ĺ–ī–į—Ä—ā–Ĺ–ĺ–ľ—É —Ā—ā–į–Ĺ—Ė (–∑–į 25 ¬įC, 100 –ļ–ü–į)
–Ü–Ĺ—Ā—ā—Ä—É–ļ—Ü—Ė—Ź –∑ –≤–ł–ļ–ĺ—Ä–ł—Ā—ā–į–Ĺ–Ĺ—Ź —ą–į–Ī–Ľ–ĺ–Ĺ—É
–ü—Ä–ł–ľ—Ė—ā–ļ–ł –ļ–į—Ä—ā–ļ–ł
–ź—Ä–≥—Ė–Ĺ—ĖŐĀ–Ĺ (őī-–≥—É–į–Ĺ—Ė–ī–ł–Ĺ-őĪ-–į–ľ—Ė–Ĺ–ĺ–≤–į–Ľ–Ķ—Ä—Ė–į–Ĺ–ĺ–≤–į –ļ–ł—Ā–Ľ–ĺ—ā–į , —Ā–ļ–ĺ—Ä–ĺ—á–Ķ–Ĺ–Ĺ—Ź –ź—Ä–≥ , Arg , R ) ‚ÄĒ –ĺ—Ā–Ĺ–ĺ–≤–Ĺ–į őĪ-–į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–į , l -—Ą–ĺ—Ä–ľ–į–≥–Ķ–Ĺ–Ķ—ā–ł—á–Ĺ–ł–ľ –ļ–ĺ–ī–ĺ–ľ (–ļ–ĺ–ī–ĺ–Ĺ–ł –¶–ď–ź, –¶–ď–£, –¶–ď–ď, –¶–ď–¶, –ź–ď–ź —ā–į –ź–ď–ď –≤ –ľ–†–Ě–ö ) —Ė —Ā—ā–į–Ĺ–ĺ–≤–Ľ—Ź—ā—Ć –ĺ—Ā–Ĺ–ĺ–≤—É –Ī—Ė–Ľ–ļ—Ė–≤. –Ē–Ľ—Ź –Ľ—é–ī–ł–Ĺ–ł –į—Ä–≥—Ė–Ĺ—Ė–Ĺ —Ē –Ĺ–į–Ņ—Ė–≤–Ĺ–Ķ–∑–į–ľ—Ė–Ĺ–Ĺ–ĺ—é –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–ĺ—é, —ā–ĺ–Ī—ā–ĺ –Ī—Ė–ĺ—Ö—Ė–ľ—Ė—á–Ĺ—Ė —ą–Ľ—Ź—Ö–ł –ī–Ľ—Ź —ó—ó –Ī—Ė–ĺ—Ā–ł–Ĺ—ā–Ķ–∑—É —Ė—Ā–Ĺ—É—é—ā—Ć, –Ņ—Ä–ĺ—ā–Ķ –≤ –Ņ–Ķ–≤–Ĺ—Ė –Ņ–Ķ—Ä—Ė–ĺ–ī–ł –∂–ł—ā—ā—Ź, –∑–ĺ–ļ—Ä–Ķ–ľ–į —Ė–Ĺ—ā–Ķ–Ĺ—Ā–ł–≤–Ĺ–ĺ–≥–ĺ —Ä–ĺ—Ā—ā—É —ā–į —Ä–ĺ–∑–≤–ł—ā–ļ—É, –į —ā–į–ļ–ĺ–∂ –Ņ—Ė–ī —á–į—Ā –ī–Ķ—Ź–ļ–ł—Ö –∑–į—Ö–≤–ĺ—Ä—é–≤–į–Ĺ—Ć –≤–ĺ–Ĺ–ł –Ĺ–Ķ –ľ–ĺ–∂—É—ā—Ć –∑–į–Ī–Ķ–∑–Ņ–Ķ—á—É–≤–į—ā–ł –ī–ĺ—Ā—ā–į—ā–Ĺ—Ć–ĺ—ó –ļ—Ė–Ľ—Ć–ļ–ĺ—Ā—ā—Ė —Ü—Ė—Ē—ó —Ā–Ņ–ĺ–Ľ—É–ļ–ł, —á–Ķ—Ä–Ķ–∑ —Č–ĺ –≤–ĺ–Ĺ–į –Ņ–ĺ–≤–ł–Ĺ–Ĺ–į –Ņ–ĺ—ā—Ä–į–Ņ–Ľ—Ź—ā–ł –≤ –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ —Ė–∑ —ó–∂–Ķ—é. –Ē–Ľ—Ź –ī–Ķ—Ź–ļ–ł—Ö –≤–ł–ī—Ė–≤ —ā–≤–į—Ä–ł–Ĺ, —Ź–ļ-–ĺ—ā –Ņ—ā–į—Ö—Ė–≤, —Ö–ł–∂–ł—Ö —ā–į –ľ–ĺ—Ä—Ā—Ć–ļ–ł—Ö —Ā—Ā–į–≤—Ü—Ė–≤, –į—Ä–≥—Ė–Ĺ—Ė–Ĺ —Ē –Ĺ–Ķ–∑–į–ľ—Ė–Ĺ–Ĺ–ĺ—é –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–ĺ—é[ 3]
–ó–į–≤–ī—Ź–ļ–ł –≥—É–į–Ĺ—Ė–ī–ł–Ĺ–ĺ–≤–ĺ–ľ—É —É–≥—Ä—É–Ņ—É–≤–į–Ĺ–Ĺ—é —Ē –Ĺ–į–Ļ–ĺ—Ā–Ĺ–ĺ–≤–Ĺ—Ė—ą–ĺ—é –∑ –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā. –ß–į—Ā—ā–ĺ –≤—Ö–ĺ–ī–ł—ā—Ć –ī–ĺ —á–į—Ā—ā–ł–Ĺ –Ī—Ė–Ľ–ļ—Ė–≤, —Č–ĺ –≤–∑–į—Ē–ľ–ĺ–ī—Ė—é—ā—Ć –∑ –Ē–Ě–ö . –£ –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ—Ė —ā–≤–į—Ä–ł–Ĺ –ī–Ķ–≥—Ä–į–ī—É—Ē—ā—Ć—Ā—Ź –ī–ĺ —Ā–Ķ—á–ĺ–≤–ł–Ĺ–ł .
–ě–ļ—Ä—Ė–ľ —ā–ĺ–≥–ĺ, —Č–ĺ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ –≤—Ö–ĺ–ī–ł—ā—Ć –ī–ĺ —Ā–ļ–Ľ–į–ī—É –Ī—Ė–Ľ–ļ—Ė–≤, —Ü—Ź –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–į —ā–į–ļ–ĺ–∂ –≤–į–∂–Ľ–ł–≤–į –ī–Ľ—Ź –Ī–į–≥–į—ā—Ć–ĺ—Ö —Ė–Ĺ—ą–ł—Ö –ľ–Ķ—ā–į–Ī–ĺ–Ľ—Ė—á–Ĺ–ł—Ö —ą–Ľ—Ź—Ö—Ė–≤ –ļ–Ľ—Ė—ā–ł–Ĺ–ł, –∑–ĺ–ļ—Ä–Ķ–ľ–į —Ā–ł–Ĺ—ā–Ķ–∑—É –Ě—Ė—ā—Ä–ĺ–≥–Ķ–Ĺ (II) –ĺ–ļ—Ā–ł–ī—É , –Ņ–ĺ–Ľ—Ė–į–ľ—Ė–Ĺ—Ė–≤ , –Ņ—Ä–ĺ–Ľ—Ė–Ĺ—É , –≥–Ľ—É—ā–į–ľ–į—ā—É , –ļ—Ä–Ķ–į—ā–ł–Ĺ—É —ā–į –į–≥–ľ–į—ā–ł–Ĺ—É [ 4]
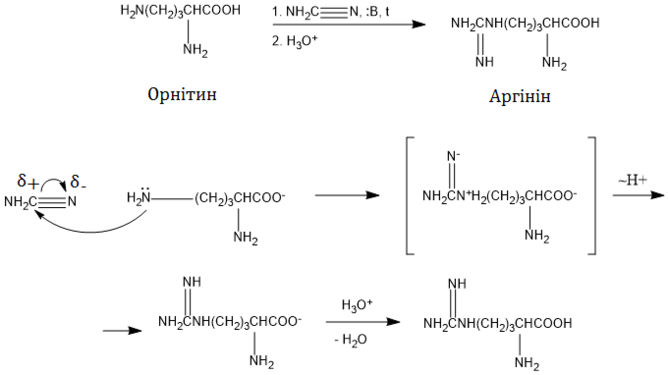
–ź—Ä–≥—Ė–Ĺ—Ė–Ĺ –Ī—É–≤ –≤–Ņ–Ķ—Ä—ą–Ķ –≤–ł–ī—Ė–Ľ–Ķ–Ĺ–ł–Ļ —Ė–∑ –Ņ—Ä–ĺ—Ä–ĺ—Ā—ā–ļ—Ė–≤ –Ľ—é–Ņ–ł–Ĺ—É 1886 —Ä–ĺ–ļ—É, –į —á–Ķ—Ä–Ķ–∑ 9 —Ä–ĺ–ļ—Ė–≤ –∑'—Ź—Ā—É–≤–į–Ľ–ĺ—Ā—Ź, —Č–ĺ –≤—Ė–Ĺ –≤—Ö–ĺ–ī–ł—ā—Ć –ī–ĺ —Ā–ļ–Ľ–į–ī—É —ā–≤–į—Ä–ł–Ĺ–Ĺ–ł—Ö –Ī—Ė–Ľ–ļ—Ė–≤. 1897 —Ä–ĺ–ļ—É –≤–ī–į–Ľ–ĺ—Ā—Ź –≤—Ā—ā–į–Ĺ–ĺ–≤–ł—ā–ł —Ö—Ė–ľ—Ė—á–Ĺ—É –Ī—É–ī–ĺ–≤—É —Ü—Ė—Ē—ó –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–ł —ą–Ľ—Ź—Ö–ĺ–ľ —ó—ó –Ľ—É–∂–Ĺ–ĺ–≥–ĺ –≥—Ė–ī—Ä–ĺ–Ľ—Ė–∑—É –ī–ĺ –ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ—É —ā–į —Ā–Ķ—á–ĺ–≤–ł–Ĺ–ł, –į –Ņ—Ė–∑–Ĺ—Ė—ą–Ķ (—É 1910 —Ä.) —Ā—ā—Ä—É–ļ—ā—É—Ä–į –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –Ī—É–Ľ–į –Ņ—Ė–ī—ā–≤–Ķ—Ä–ī–∂–Ķ–Ĺ–į —ą–Ľ—Ź—Ö–ĺ–ľ –Ļ–ĺ–≥–ĺ —Ā–ł–Ĺ—ā–Ķ–∑—É —Ė–∑ –Ī–Ķ–Ĺ–∑–ł–Ľ–ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ—É . 1924 –Ī—É–Ľ–ĺ –≤—Ė–ī–ļ—Ä–ł—ā–ĺ, —Č–ĺ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ —Ē –≥–ĺ–Ľ–ĺ–≤–Ĺ–ĺ—é –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–ĺ—é –≤ –ĺ—Ā–Ĺ–ĺ–≤–Ĺ–ł—Ö –Ī—Ė–Ľ–ļ–į—Ö —Ā–Ņ–Ķ—Ä–ľ–ł —Ä–ł–Ī. –ź 1930 —Ä–ĺ—Ü—Ė –í—Ė–Ľ—Ć—Ź–ľ –ö–į–ľ–ľ—Ė–Ĺ–≥ –†–ĺ—É–∑ –Ņ–ĺ–ļ–į–∑–į–≤ —É —Ā–≤–ĺ—Ē–ľ—É –ļ–Ľ–į—Ā–ł—á–Ĺ–ĺ–ľ—É –Ķ–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā—Ė, —Č–ĺ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ –Ĺ–į–Ľ–Ķ–∂–ł—ā—Ć –ī–ĺ –∑–į–ľ—Ė–Ĺ–Ĺ–ł—Ö –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā, —ā–ĺ–Ī—ā–ĺ –ľ–ĺ–∂–Ķ —Ā–ł–Ĺ—ā–Ķ–∑—É–≤–į—ā–ł—Ā—Ź –≤ –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ—Ė —Ā—Ā–į–≤—Ü—Ė–≤ . 1932 —Ä–ĺ–ļ—É –ď–į–Ĺ—Ā –ö—Ä–Ķ–Ī—Ā —ā–į –ö—É—Ä—ā –ď–Ķ–Ĺ–∑–Ķ–Ľ—Ź–Ļ—ā –≤—Ė–ī–ļ—Ä–ł–Ľ–ł —Ü–ł–ļ–Ľ —Ā–Ķ—á–ĺ–≤–ł–Ĺ–ł , —É —Ź–ļ–ĺ–ľ—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ —Ē –Ņ—Ä–ĺ–ľ—Ė–∂–Ĺ–ł–ľ –ľ–Ķ—ā–į–Ī–ĺ–Ľ—Ė—ā–ĺ–ľ, —Č–ĺ –Ņ–ĺ–ļ–Ľ–į–Ľ–ĺ –Ņ–ĺ—á–į—ā–ĺ–ļ –ī–Ķ—ā–į–Ľ—Ć–Ĺ–ĺ–ľ—É –≤–ł–≤—á–Ķ–Ĺ–Ĺ—é –ľ–Ķ—ā–į–Ī–ĺ–Ľ—Ė–∑–ľ—É —Ü—Ė—Ē—ó –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–ł[ 5]
–£ –Ņ—Ė–∑–Ĺ—Ė 1930-—ā—Ė —ā–į 1940-–≤—Ė —Ä–ĺ–ļ–ł –§–ĺ—Ā—ā–Ķ—Ä –≤—Ā—ā–į–Ĺ–ĺ–≤–ł–≤, —Č–ĺ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ —Ē –Ņ–ĺ–Ņ–Ķ—Ä–Ķ–ī–Ĺ–ł–ļ–ĺ–ľ —É —Ā–ł–Ĺ—ā–Ķ–∑—Ė –ļ—Ä–Ķ–į—ā–ł–Ĺ—É. –Ę–ĺ–ī—Ė –∂ –Ī—É–Ľ–ĺ –≤–ł—Ź–≤–Ľ–Ķ–Ĺ–ĺ, —Č–ĺ –Ĺ–į–ī—Ö–ĺ–ī–∂–Ķ–Ĺ–Ĺ—Ź —Ü—Ė—Ē—ó –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–ł —Ė–∑ –ī—Ė—Ē—ā–ĺ—é –Ĺ–Ķ–ĺ–Ī—Ö—Ė–ī–Ĺ–Ķ –ī–Ľ—Ź —Ä–ĺ—Ā—ā—É –ļ—É—Ä—á–į—ā —Ė –ľ–ĺ–Ľ–ĺ–ī–ł—Ö –Ņ–į—Ü—é–ļ—Ė–≤ , –į–Ľ–Ķ –Ĺ–Ķ –ĺ–Ī–ĺ–≤'—Ź–∑–ļ–ĺ–≤–Ķ –ī–Ľ—Ź –Ĺ–ĺ—Ä–ľ–į–Ľ—Ć–Ĺ–ĺ—ó –∂–ł—ā—ā—Ē–ī—Ė—Ź–Ľ—Ć–Ĺ–ĺ—Ā—ā—Ė –ī–ĺ—Ä–ĺ—Ā–Ľ–ł—Ö –∑–ī–ĺ—Ä–ĺ–≤–ł—Ö –Ņ–į—Ü—é–ļ—Ė–≤. –ü–ĺ–ī–į–Ľ—Ć—ą—Ė –ī–ĺ—Ā–Ľ—Ė–ī–∂–Ķ–Ĺ–Ĺ—Ź, —Č–ĺ —ā—Ä–ł–≤–į–Ľ–ł –ī–ĺ 1970-—Ö —Ä–ĺ–ļ—Ė–≤, –Ņ–ĺ–ļ–į–∑–į–Ľ–ł, —Č–ĺ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ ‚ÄĒ –∑–į–ľ—Ė–Ĺ–Ĺ–į –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–į –ī–Ľ—Ź –ī–ĺ—Ä–ĺ—Ā–Ľ–ł—Ö –∑–ī–ĺ—Ä–ĺ–≤–ł—Ö –Ľ—é–ī–Ķ–Ļ —ā–į –Ī–į–≥–į—ā—Ć–ĺ—Ö —Ė–Ĺ—ą–ł—Ö —Ā—Ā–į–≤—Ü—Ė–≤, –į–Ľ–Ķ –Ĺ–Ķ–∑–į–ľ—Ė–Ĺ–Ĺ–į –ī–Ľ—Ź —ó—Ö–Ĺ—Ė—Ö –ľ–į–Ľ—Ź—ā —ā–į –ī–Ľ—Ź —Ö–ł–∂–į–ļ—Ė–≤ [ 5]
–í—Ė–Ĺ–ī–ľ—é–Ľ–Ľ–Ķ—Ä (–į–Ĺ–≥–Ľ. Windmueller ) —ā–į –°–Ņ–į—Ē—ā (–į–Ĺ–≥–Ľ. Spaeth ) —É 1980-—Ö —Ä–ĺ–ļ–į—Ö –Ņ–ĺ–ļ–į–∑–į–Ľ–ł, —Č–ĺ –≥–ĺ–Ľ–ĺ–≤–Ĺ–ł–ľ –ī–∂–Ķ—Ä–Ķ–Ľ–ĺ–ľ —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ—É –≤ –ļ—Ä–ĺ–≤—Ė, —Ź–ļ–ł–Ļ –≤–ł–ļ–ĺ—Ä–ł—Ā—ā–ĺ–≤—É—Ē—ā—Ć—Ā—Ź —ā–ļ–į–Ĺ–ł–Ĺ–į–ľ–ł –ī–Ľ—Ź —Ā–ł–Ĺ—ā–Ķ–∑—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É, —Ē —ā–ĺ–Ĺ–ļ–į –ļ–ł—ą–ļ–į . –ó–≥–ĺ–ī–ĺ–ľ —Ü–Ķ –Ņ—Ä–ł–∑–≤–Ķ–Ľ–ĺ –ī–ĺ –≤—Ė–ī–ļ—Ä–ł—ā—ā—Ź –ľ–Ķ—ā–į–Ī–ĺ–Ľ—Ė—á–Ĺ–ĺ–≥–ĺ —ą–Ľ—Ź—Ö—É –Ī—Ė–ĺ—Ā–ł–Ĺ—ā–Ķ–∑—É —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ—É —Ė–∑ –≥–Ľ—É—ā–į–ľ—Ė–Ĺ—É/–≥–Ľ—É—ā–į–ľ–į—ā—É —á–Ķ—Ä–Ķ–∑ őĒ1 -–Ņ—Ė—Ä–ĺ–Ľ—Ė–Ĺ-5-–ļ–į—Ä–Ī–ĺ–ļ—Ā–ł–Ľ–į—ā. –Ę–į–ļ–ĺ–∂ –Ĺ–į–Ņ—Ä–ł–ļ—Ė–Ĺ—Ü—Ė 1980-—Ö —Ä–ĺ–ļ—Ė–≤ –Ī—É–Ľ–į –∑'—Ź—Ā–ĺ–≤–į–Ĺ–į —Ä–ĺ–Ľ—Ć –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É —Ź–ļ –Ņ–ĺ–Ņ–Ķ—Ä–Ķ–ī–Ĺ–ł–ļ–į —É —Ā–ł–Ĺ—ā–Ķ–∑—Ė NO –≤ –Ķ–ī–ĺ—ā–Ķ–Ľ—Ė—ó —ā–į –ľ–į–ļ—Ä–ĺ—Ą–į–≥–į—Ö , –į –Ņ—Ė–∑–Ĺ—Ė—ą–Ķ —Ė –≤ –Ī–į–≥–į—ā—Ć–ĺ—Ö —Ė–Ĺ—ą–ł—Ö —ā–ł–Ņ–į—Ö –ļ–Ľ—Ė—ā–ł–Ĺ[ 5]
–Ē–Ķ–Ľ–ĺ–ļ–į–Ľ—Ė–∑–į—Ü—Ė—Ź –Ņ–ĺ–∑–ł—ā–ł–≤–Ĺ–ĺ–≥–ĺ –∑–į—Ä—Ź–ī—É –≤ –≥—É–į–Ĺ—Ė–ī–ł–Ĺ–ĺ–≤–ĺ–ľ—É —É–≥—Ä—É–Ņ—É–≤–į–Ĺ–Ĺ—Ė l -–į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –ź—Ä–≥—Ė–Ĺ—Ė–Ĺ —Ź–ļ őĪ-–į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–į –ľ—Ė—Ā—ā–ł—ā—Ć –į–ľ—Ė–Ĺ–ĺ- —ā–į –ļ–į—Ä–Ī–ĺ–ļ—Ā–ł–Ľ—Ć–Ĺ—É –≥—Ä—É–Ņ—É —Ä–ĺ–∑—ā–į—ą–ĺ–≤–į–Ĺ—Ė –Ī—Ė–Ľ—Ź –ĺ–ī–Ĺ–ĺ–≥–ĺ (őĪ) –į—ā–ĺ–ľ–į –ö–į—Ä–Ī–ĺ–Ĺ—É . –¶–Ķ–Ļ –į—ā–ĺ–ľ —Ē —Ö—Ė—Ä–į–Ľ—Ć–Ĺ–ł–ľ, —Ė –≤ –∑–į–Ľ–Ķ–∂–Ĺ–ĺ—Ā—ā—Ė –≤—Ė–ī —Ä–ĺ–∑—ā–į—ą—É–≤–į–Ĺ–Ĺ—Ź –∑–į–ľ—Ė—Ā–Ĺ–ł–ļ—Ė–≤ –Ī—Ė–Ľ—Ź –Ĺ—Ć–ĺ–≥–ĺ —Ä–ĺ–∑—Ä—Ė–∑–Ĺ—Ź—é—ā—Ć l - —ā–į d -—Ą–ĺ—Ä–ľ–ł, –ī–ĺ —Ā–ļ–Ľ–į–ī—É –Ī—Ė–Ľ–ļ—Ė–≤ –≤—Ö–ĺ–ī–ł—ā—Ć –Ņ–Ķ—Ä—ą–į —Ė–∑ –Ĺ–ł—Ö.
R-–≥—Ä—É–Ņ–į –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É —Ā–ļ–Ľ–į–ī–į—Ē—ā—Ć—Ā—Ź —Ė–∑ –ļ–į—Ä–Ī–ĺ–Ĺ–ĺ–≤–ĺ–≥–ĺ –Ľ–į–Ĺ—Ü—é–≥–į –ī–ĺ–≤–∂–ł–Ĺ–ĺ—é —ā—Ä–ł –į—ā–ĺ–ľ–ł, –Ĺ–į –ļ—Ė–Ĺ—Ü—Ė —Ź–ļ–ĺ–≥–ĺ —Ä–ĺ–∑—ā–į—ą–ĺ–≤–į–Ĺ–Ķ –≥—É–į–Ĺ—Ė–ī–ł–Ĺ–ĺ–≤–Ķ —É–≥—Ä—É–Ņ—É–≤–į–Ĺ–Ĺ—Ź, –∑–į–≤–ī—Ź–ļ–ł —Ź–ļ–ĺ–ľ—É —Ü—Ź –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–į –ľ–į—Ē —Ā–ł–Ľ—Ć–Ĺ—Ė –ĺ—Ā–Ĺ–ĺ–≤–Ĺ—Ė –≤–Ľ–į—Ā—ā–ł–≤–ĺ—Ā—ā—Ė. –ö–ĺ–Ľ–ł –į—Ä–≥—Ė–Ĺ—Ė–Ĺ –Ņ–Ķ—Ä–Ķ–Ī—É–≤–į—Ē –≤ —Ė–ĺ–Ĺ—Ė–∑–ĺ–≤–į–Ĺ—Ė–Ļ —Ą–ĺ—Ä–ľ—Ė, –Ņ–ĺ–∑–ł—ā–ł–≤–Ĺ–ł–Ļ –∑–į—Ä—Ź–ī —É –≥—É–į–Ĺ—Ė–ī–ł–Ĺ–ĺ–≤—Ė–Ļ –≥—Ä—É–Ņ—Ė —Ä—Ė–≤–Ĺ–ĺ–ľ—Ė—Ä–Ĺ–ĺ —Ä–ĺ–∑–Ņ–ĺ–ī—Ė–Ľ—Ź—Ē—ā—Ć—Ā—Ź –ľ—Ė–∂ —ā—Ä—Ć–ĺ–ľ–į –į—ā–ĺ–ľ–į–ľ–ł –Ě—Ė—ā—Ä–ĺ–≥–Ķ–Ĺ—É –Ļ –į—ā–ĺ–ľ–ĺ–ľ –ö–į—Ä–Ī–ĺ–Ĺ—É –∑–į–≤–ī—Ź–ļ–ł –ī–Ķ–Ľ–ĺ–ļ–į–Ľ—Ė–∑–į—Ü—Ė—ó –Ķ–Ľ–Ķ–ļ—ā—Ä–ĺ–Ĺ—Ė–≤ .
–õ–ĺ–≥–į—Ä–ł—Ą–ľ—Ė—á–Ĺ–į –ļ–ĺ–Ĺ—Ā—ā–į–Ĺ—ā–į –ī–ł—Ā–ĺ—Ü—Ė–į—Ü—Ė—ó –ļ–į—Ä–Ī–ĺ–ļ—Ā–ł–Ľ—Ć–Ĺ–ĺ—ó –≥—Ä—É–Ņ–ł –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É pK1 = 2,17; –ļ–ĺ–Ĺ—Ā—ā–į–Ĺ—ā–į –ī–ł—Ā–ĺ—Ü—Ė–į—Ü—Ė—ó őĪ-–į–ľ—Ė–Ĺ–ĺ–≥—Ä—É–Ņ–ł pK2 = 9,04, –į –≥—É–į–Ĺ—Ė–ī–ł–Ĺ–≤–ĺ–≥–ĺ —É–≥—Ä—É–Ņ—É–≤–į–Ĺ–Ĺ—Ź pKR (R –≥—Ä—É–Ņ–į) = 12,48. –ź—Ä–≥—Ė–Ĺ—Ė–Ĺ –Ĺ–į–Ļ–ĺ—Ā–Ĺ–ĺ–≤–Ĺ—Ė—ą–į —Ė–∑ 20 —Ā—ā–į–Ĺ–ī–į—Ä—ā–Ĺ–ł—Ö –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā, —Ė–∑–ĺ–Ķ–Ľ–Ķ–ļ—ā—Ä–ł—á–Ĺ–į —ā–ĺ—á–ļ–į –ī–Ľ—Ź –Ĺ–Ķ—ó —Ā—ā–į–Ĺ–ĺ–≤–ł—ā—Ć pI = 10,76.
–Ü–Ĺ–ī–Ķ–ļ—Ā –≥—Ė–ī—Ä–ĺ–Ņ–į—ā–ł—á–Ĺ–ĺ—Ā—ā—Ė (–∑–į–ľ—Ė–Ĺ–į –≤—Ė–Ľ—Ć–Ĺ–ĺ—ó –Ķ–Ĺ–Ķ—Ä–≥—Ė—ó –Ņ—Ä–ł –Ņ–Ķ—Ä–Ķ–Ĺ–Ķ—Ā–Ķ–Ĺ–Ĺ—Ė R –≥—Ä—É–Ņ–ł —Ė–∑ –≥—Ė–ī—Ä–ĺ—Ą–ĺ–Ī–Ĺ–ĺ–≥–ĺ —Ä–ĺ–∑—á–ł–Ĺ–Ĺ–ł–ļ–į —É –≤–ĺ–ī—É) –ī–Ľ—Ź –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É —Ā—ā–į–Ĺ–ĺ–≤–ł—ā—Ć -4,5 –ļ–Ē–∂/–ľ–ĺ–Ľ—Ć, –≤–ĺ–Ĺ–į –Ĺ–į–Ļ–Ī—Ė–Ľ—Ć—ą –≥—Ė–ī—Ä–ĺ—Ą—Ė–Ľ—Ć–Ĺ–į —Ė–∑ 20 —Ā—ā–į–Ĺ–ī–į—Ä—ā–Ĺ–ł—Ö –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā.
–£ —Ä–ĺ–∑—á–ł–Ĺ—Ė –į–Ī–ĺ —É —Ā–ļ–Ľ–į–ī—Ė –Ī—Ė–Ľ–ļ—Ė–≤ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ –ľ–ĺ–∂–Ĺ–į —Ź–ļ—Ė—Ā–Ĺ–ĺ —ā–į –ļ—Ė–Ľ—Ć–ļ—Ė—Ā–Ĺ–ĺ –≤–ł–∑–Ĺ–į—á–ł—ā–ł –∑–į–≤–ī—Ź–ļ–ł —Ä–Ķ–į–ļ—Ü—Ė—ó –°–į–ļ–į–≥—É—á—Ė . –í–ĺ–Ĺ–į –Ņ–ĺ–Ľ—Ź–≥–į—Ē —É –≤–∑–į—Ē–ľ–ĺ–ī—Ė—ó –≥—Ė–Ņ–ĺ—Ö–Ľ–ĺ—Ä–ł—ā—É –Ĺ–į—ā—Ä—Ė—é —Ė 2-–Ĺ–į—Ą—ā–ĺ–Ľ—É –∑ –≥—É–į–Ĺ—Ė–ī–ł–Ĺ–ĺ–ľ —É –Ľ—É–∂–Ĺ–ĺ–ľ—É —Ā–Ķ—Ä–Ķ–ī–ĺ–≤–ł—Č—Ė –∑ —É—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź–ľ —Ā–Ņ–ĺ–Ľ—É–ļ —á–Ķ—Ä–≤–ĺ–Ĺ–ĺ–≥–ĺ –∑–į–Ī–į—Ä–≤–Ľ–Ķ–Ĺ–Ĺ—Ź[ 9]
–ě–ī–Ķ—Ä–∂–į–Ĺ–Ĺ—Ź –ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ—É
–ě—Ā–Ĺ–ĺ–≤–į: –í –Ņ–ĺ–≤'—Ź–∑—É—Ē –Ņ—Ä–ĺ—ā–ĺ–Ĺ –ļ–į—Ä–Ī–ĺ–ļ—Ā–ł–Ľ—Ć–Ĺ–ĺ—ó –≥—Ä—É–Ņ–ł, –Ņ—Ė—Ā–Ľ—Ź —á–ĺ–≥–ĺ —Ā–Ľ—Ė–ī—É—Ē –Ĺ—É–ļ–Ľ–Ķ–ĺ—Ą—Ė–Ľ—Ć–Ĺ–į –į—ā–į–ļ–į őī-NH2 -–≥—Ä—É–Ņ–ł (–Ī—Ė–Ľ—Ć—ą —Ā–ł–Ľ—Ć–Ĺ–ł–Ļ –Ĺ—É–ļ–Ľ–Ķ–ĺ—Ą—Ė–Ľ—Ć–Ĺ–ł–Ļ —Ü–Ķ–Ĺ—ā—Ä, –Ĺ—Ė–∂ őĪ-–į–ľ—Ė–Ĺ–ĺ–≥—Ä—É–Ņ–į) –Ņ–ĺ
C
≡ ‚Č° -->
N
{\displaystyle C\equiv N}
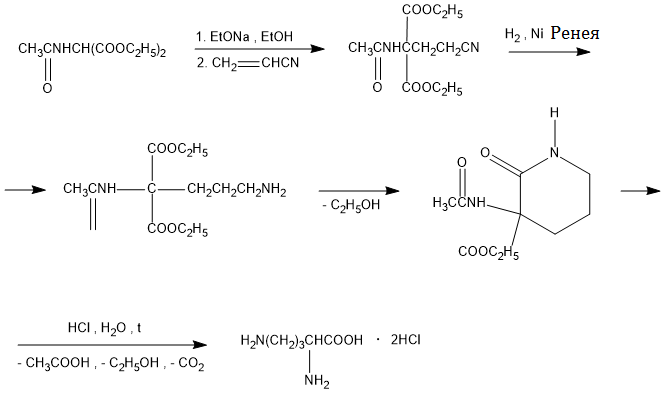
–Ď—Ė–ĺ—Ā–ł–Ĺ—ā–Ķ–∑ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≤ —Ā—Ā–į–≤—Ü—Ė–≤
–ě—Ā–Ĺ–ĺ–≤–Ĺ–ł–ľ–ł –ī–∂–Ķ—Ä–Ķ–Ľ–į–ľ–ł –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≤ –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ—Ė –Ľ—é–ī–ł–Ĺ–ł —Ē —ó–∂–į, –ĺ–Ī–ľ—Ė–Ĺ –Ī—Ė–Ľ–ļ—Ė–≤ —ā–į –Ķ–Ĺ–ī–ĺ–≥–Ķ–Ĺ–Ĺ–ł–Ļ —Ā–ł–Ĺ—ā–Ķ–∑. –ü—Ä–ł —á–ĺ–ľ—É —É –ī–ĺ—Ä–ĺ—Ā–Ľ–ł—Ö –ĺ—Ā—Ė–Ī —ā—Ė–Ľ—Ć–ļ–ł 5‚ÄĒ15 % –∑–į–≥–į–Ľ—Ć–Ĺ–ĺ—ó –ļ—Ė–Ľ—Ć–ļ–ĺ—Ā—ā—Ė —Ü—Ė—Ē—ó –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–ł –≤ –ļ—Ä–ĺ–≤—Ė –Ņ—Ä–ł–Ņ–į–ī–į—Ē –Ĺ–į —É—ā–≤–ĺ—Ä–Ķ–Ĺ—É de novo . –Ď—Ė–Ľ—Ć—ą—Ė—Ā—ā—Ć —Ā–ł–Ĺ—ā–Ķ–∑—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≤ –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ—Ė –≤—Ė–ī–Ī—É–≤–į—Ē—ā—Ć—Ā—Ź –∑–į —É—á–į—Ā—ā—Ė —ā–ĺ–Ĺ–ļ–ĺ–≥–ĺ –ļ–ł—ą–Ķ—á–Ĺ–ł–ļ–į —ā–į –Ĺ–ł—Ä–ĺ–ļ [ 5]
–Ď—Ė–ĺ—Ā–ł–Ĺ—ā–Ķ–∑ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É de novo —Ä–ĺ–∑–Ņ–ĺ—á–ł–Ĺ–į—Ē—ā—Ć—Ā—Ź —Ė–∑ –≥–Ľ—É—ā–į–ľ–į—ā—É , —Ź–ļ–ł–Ļ —Ą–ĺ—Ā—Ą–ĺ—Ä–ł–Ľ—é—Ē—ā—Ć—Ā—Ź –ī–ĺ ő≥-–≥–Ľ—É—ā–į–ľ—Ė–Ľ—Ą–ĺ—Ā—Ą–į—ā—É —Ė–∑ –≤–ł–ļ–ĺ—Ä–ł—Ā—ā–į–Ĺ–Ĺ—Ź–ľ –ĺ–ī–Ĺ—Ė—Ē—ó –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–ł –ź–Ę–§ , –Ņ—Ė—Ā–Ľ—Ź —á–ĺ–≥–ĺ —É—ā–≤–ĺ—Ä–Ķ–Ĺ–ł–Ļ –Ņ—Ä–ĺ–ī—É–ļ—ā –≤—Ė–ī–Ĺ–ĺ–≤–Ľ—é—Ē—ā—Ć—Ā—Ź –ī–ĺ –≥–Ľ—É—ā–į–ľ–į—ā-ő≥-—Ā–Ķ–ľ—Ė–į–ī—Ć–ī–Ķ–≥—Ė–ī—É . –¶—Ė —Ä–Ķ–į–ļ—Ü—Ė—ó –ļ–į—ā–į–Ľ—Ė–∑—É—é—ā—Ć –≥–Ľ—É—ā–į–ľ–į—ā–ļ—Ė–Ĺ–į–∑–į —ā–į ő≥-–≥–Ľ—É—ā–į–ľ—Ė–Ľ—Ą–ĺ—Ā—Ą–į—ā—Ä–Ķ–ī—É–ļ—ā–į–∑–į –≤—Ė–ī–Ņ–ĺ–≤—Ė–ī–Ĺ–ĺ. –ď–Ľ—É—ā–į–ľ–į—ā-ő≥-—Ā–Ķ–ľ—Ė–į–Ľ—Ć–ī–Ķ–≥—Ė–ī —Ā–Ņ–ĺ–Ĺ—ā–į–Ĺ–Ĺ–ĺ —Ü–ł–ļ–Ľ—Ė–∑—É—Ē—ā—Ć—Ā—Ź —É őĒ1 -–Ņ—Ė—Ä–ĺ–Ľ—Ė–Ĺ-5-–ļ–į—Ä–Ī–ĺ–ļ—Ā–ł–Ľ–į—ā (–ü5–ö), —Ź–ļ–ł–Ļ –ī–į–Ľ—Ė –ľ–ĺ–∂–Ķ –Ī—É—ā–ł –Ņ–Ķ—Ä–Ķ—ā–≤–ĺ—Ä–Ķ–Ĺ–ł–Ļ —É –Ņ—Ä–ĺ–Ľ—Ė–Ĺ , –Ņ—Ä–ĺ—ā–Ķ –Ņ—Ė–ī —á–į—Ā –Ī—Ė–ĺ—Ā–ł–Ĺ—ā–Ķ–∑—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≥–Ľ—É—ā–į–ľ–į—ā-ő≥-—Ā–Ķ–ľ—Ė–į–Ľ—Ć–ī–Ķ–≥—Ė–ī –Ņ–ĺ–≤–ł–Ĺ–Ķ–Ĺ –≤—Ā—ā—É–Ņ–į—ā–ł –≤ —Ä–Ķ–į–ļ—Ü—Ė—é –Ņ–Ķ—Ä–Ķ–į–ľ—Ė–Ĺ—É–≤–į–Ĺ–Ĺ—Ź —Ė–∑ –≥–Ľ—É—ā–į–ľ–į—ā–ĺ–ľ, –≤–Ĺ–į—Ā–Ľ—Ė–ī–ĺ–ļ —Ź–ļ–ĺ—ó –≤—Ė–Ĺ –Ņ–Ķ—Ä–Ķ—ā–≤–ĺ—Ä—é—Ē—ā—Ć—Ā—Ź –ī–ĺ l -–ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ—É . –†—Ė–≤–Ĺ–ĺ–≤–į–≥–į —Ä–Ķ–į–ļ—Ü—Ė—ó —Ü–ł–ļ–Ľ—Ė–∑–į—Ü—Ė—ó —Ā–Ķ–ľ—Ė–į–Ľ—Ć–ī–Ķ–≥—É —É –ü5–ö —Ā–Ņ—Ä–ł—Ź—Ē —É—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—é –ĺ—Ā—ā–į–Ĺ–Ĺ—Ć–ĺ–≥–ĺ, –Ņ—Ä–ĺ—ā–Ķ –≤–ĺ–Ĺ–į –ľ–ĺ–∂–Ķ –Ņ—Ä–ĺ—ā—Ė–ļ–į—ā–ł —Ė –≤ –Ĺ–į–Ņ—Ä—Ź–ľ–ļ—É –Ĺ–Ķ–ĺ–Ī—Ö—Ė–ī–Ĺ–ĺ–ľ—É –ī–Ľ—Ź —Ā–ł–Ĺ—ā–Ķ–∑—É –ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ—É, –∑–į —É–ľ–ĺ–≤ –Ĺ–Ķ—Ā—ā–į—á—Ė –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≤ –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ—Ė. –ě—Ä–Ĺ—Ė—ā–ł–Ĺ —ā–į–ļ–ĺ–∂ –ľ–ĺ–∂–Ķ –Ī—É—ā–ł —Ā–ł–Ĺ—ā–Ķ–∑–ĺ–≤–į–Ĺ–ł–Ļ —Ė–∑ –Ņ—Ä–ĺ–Ľ—Ė–Ĺ—É, —Č–ĺ –ĺ–ļ–ł—Ā–Ĺ—é—Ē—ā—Ć—Ā—Ź –ī–ĺ –ü5–ö –Ņ—Ä–ĺ–Ľ—Ė–Ĺ–ĺ–ļ—Ā–ł–ī–į–∑–ĺ—é [ 5]
–ě—Ä–Ĺ—Ė—ā–ł–Ĺ —Ē –Ņ—Ä–ĺ–ľ—Ė–∂–Ĺ–ł–ľ –Ņ—Ä–ĺ–ī—É–ļ—ā–ĺ–ľ —Ü–ł–ļ–Ľ—É —Ā–Ķ—á–ĺ–≤–ł–Ĺ–ł, –≤ —Ä–Ķ–į–ļ—Ü—Ė—Ź—Ö —Ź–ļ–ĺ–≥–ĺ –≤—Ė–Ĺ –Ņ–Ķ—Ä–Ķ—ā–≤–ĺ—Ä—é—Ē—ā—Ć—Ā—Ź –ī–ĺ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É. –°–Ņ–Ķ—Ä—ą—É —Ü—Ź –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–į –≤—Ā—ā—É–Ņ–į—Ē –≤ —Ä–Ķ–į–ļ—Ü—Ė—é –ļ–ĺ–Ĺ–ī–Ķ–Ĺ—Ā–į—Ü—Ė—ó —Ė–∑ –ļ–į—Ä–Ī–į–ľ–ĺ—ó–Ľ—Ą–ĺ—Ā—Ą–į—ā–ĺ–ľ , –Ņ—Ä–ĺ–ī—É–ļ—ā–ĺ–ľ —Ź–ļ–ĺ—ó —Ē l -—Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ . –ö–į—ā–į–Ľ—Ė–∑—É—Ē —Ü–Ķ –Ņ–Ķ—Ä–Ķ—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź –ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ–ļ–į—Ä–Ī–į–ľ–ĺ—ó–Ľ—ā—Ä–į–Ĺ—Ā—Ą–Ķ—Ä–į–∑–į (—Ė–Ĺ—ą–į –Ĺ–į–∑–≤–į –ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ—ā—Ä–į–Ĺ—Ā–ļ–į—Ä–Ī–į–ľ–ĺ—ó–Ľ–į–∑–į). –ö–į—Ä–Ī–į–ľ–ĺ—ó–Ľ—Ą–ĺ—Ā—Ą–į—ā —Ā–ł–Ĺ—ā–Ķ–∑—É—Ē—ā—Ć—Ā—Ź —É –ľ—Ė—ā–ĺ—Ö–ĺ–Ĺ–ī—Ä—Ė—Ź—Ö —Ė–∑ –≥—Ė–ī—Ä–ĺ–ļ–į—Ä–Ī–ĺ–Ĺ–į—ā—É —ā–į –į–ľ—Ė–į–ļ—É —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā–ĺ–ľ –ļ–į—Ä–Ī–į–ľ–ĺ—ó–Ľ—Ą–ĺ—Ā—Ą–į—ā—Ā–ł–Ĺ—ā–į–∑–ĺ—é I , —Ź–ļ–į –Ņ–ĺ—ā—Ä–Ķ–Ī—É—Ē –Ĺ–į—Ź–≤–Ĺ–ĺ—Ā—ā—Ė N-–į—Ü–Ķ—ā–ł–Ľ–≥–Ľ—É—ā–į–ľ–į—ā—É —Ź–ļ –į–Ľ–ĺ—Ā—ā–Ķ—Ä–ł—á–Ĺ–ĺ–≥–ĺ –į–ļ—ā–ł–≤–į—ā–ĺ—Ä–į. –£ —Ü–ł—ā–ĺ–Ņ–Ľ–į–∑–ľ—Ė –ļ–Ľ—Ė—ā–ł–Ĺ–ł –į—Ä–≥—Ė–Ĺ—Ė–Ĺ–ĺ—Ā—É–ļ—Ü–ł–Ĺ–į—ā—Ā–ł–Ĺ—ā–į–∑–į (–ź–°–°) –ļ–į—ā–į–Ľ—Ė–∑—É—Ē –≤–∑–į—Ē–ľ–ĺ–ī—Ė—é —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ—É —Ė–∑ –į—Ā–Ņ–į—Ä—ā–į—ā–ĺ–ľ , –Ņ—Ä–ĺ–ī—É–ļ—ā–ĺ–ľ —Ź–ļ–ĺ—ó —Ē –į—Ä–≥—Ė–Ĺ—Ė–Ĺ–ĺ—Ā—É–ļ—Ü–ł–Ĺ–į—ā , —Č–ĺ –≤—Ė–ī—Ä–į–∑—É –∂ —Ä–ĺ–∑—Č–Ķ–Ņ–Ľ—é—Ē—ā—Ć—Ā—Ź –į—Ä–≥—Ė–Ĺ—Ė–Ĺ–ĺ—Ā—É–ļ—Ü–ł–Ĺ–į–∑–ĺ—é (–į—Ä–≥—Ė–Ĺ—Ė–Ĺ–ĺ—Ā—É–ļ—Ü–ł–Ĺ–į—ā–Ľ—Ė–į–∑–ĺ—é, –ź–°–õ) –ī–ĺ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É —ā–į —Ą—É–ľ–į—Ä–į—ā—É [ 5]
–ē–ļ—Ā–Ņ—Ä–Ķ—Ā—Ė—Ź —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā—Ė–≤ –Ī—Ė–ĺ—Ā–ł–Ĺ—ā–Ķ–∑—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –ĺ—Ä–≥–į–Ĺ–į–ľ–ł —Ā—Ā–į–≤—Ü—Ė–≤
–ě—Ä–Ĺ—Ė—ā–ł–Ĺ–į–ľ—Ė–Ĺ–ĺ—ā—Ä–į–Ĺ—Ā—Ą–Ķ—Ä–į–∑–į
–Ď—Ė–Ľ—Ć—ą—Ė—Ā—ā—Ć –ĺ—Ä–≥–į–Ĺ—Ė–≤
–ź—Ä–≥—Ė–Ĺ—Ė–Ĺ–ĺ—Ā—É–ļ—Ü–ł–Ĺ–į—ā—Ā–ł–Ĺ—ā–į–∑–į (–ź–°–°)
–ź—Ä–≥—Ė–Ĺ—Ė–Ĺ–ĺ—Ā—É–ļ—Ü–ł–Ĺ–į—ā–Ľ—Ė–į–∑–į (–ź–°–õ)
–ö–į—Ä–Ī–į–ľ–ĺ—ó–Ľ—Ą–ĺ—Ā—Ą–į—ā—Ā–ł–Ĺ—ā–į–∑–į I
–ü–Ķ—á—Ė–Ĺ–ļ–į, —ā–ĺ–Ĺ–ļ–į –ļ–ł—ą–ļ–į
–ě—Ä–Ĺ—Ė—ā–ł–Ĺ—ā—Ä–į–Ĺ—Ā–ļ–į—Ä–Ī–į–ľ–ĺ—ó–Ľ–į–∑–į
N-–į—Ü–Ķ—ā–ł–Ľ–≥–Ľ—É—ā–į–ľ–į—ā—Ā–ł–Ĺ—ā–į–∑–į
–ü—Ä–ĺ–Ľ—Ė–Ĺ–ĺ–ļ—Ā–ł–ī–į–∑–į
–ü–Ķ—á—Ė–Ĺ–ļ–į, —ā–ĺ–Ĺ–ļ–į –ļ–ł—ą–ļ–į, –Ĺ–ł—Ä–ļ–ł
–ď–Ľ—É—ā–į–ľ–į—ā–ļ—Ė–Ĺ–į–∑–į
–Ę–ĺ–Ĺ–ļ–į –ļ–ł—ą–ļ–į
ő≥-–≥–Ľ—É—ā–į–ľ—Ė–Ľ—Ą–ĺ—Ā—Ą–į—ā—Ä–Ķ–ī—É–ļ—ā–į–∑–į
–ē–ļ—Ā–Ņ—Ä–Ķ—Ā—Ė—Ź —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā—Ė–≤ —ą–Ľ—Ź—Ö—É –Ī—Ė–ĺ—Ā–ł–Ĺ—ā–Ķ–∑—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É —Ė–∑ –≥–Ľ—É—ā–į–ľ–į—ā—É –≤—Ė–ī—Ä—Ė–∑–Ĺ—Ź—Ē—ā—Ć—Ā—Ź –≤ —Ä—Ė–∑–Ĺ–ł—Ö –ĺ—Ä–≥–į–Ĺ–į—Ö. –Ē–Ķ—Ź–ļ—Ė —Ė–∑ –Ĺ–ł—Ö, —ā–į–ļ—Ė —Ź–ļ –ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ-őī-–į–ľ—Ė–Ĺ–ĺ—ā—Ä–į–Ĺ—Ā—Ą–Ķ—Ä–į–∑–į, –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—Ā—É–ļ—Ü–ł–Ĺ–į—ā—Ā–ł–Ĺ—ā–į–∑–į —ā–į –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—Ā—É–ļ—Ü–ł–Ĺ–į–∑–į, –Ĺ–į—Ź–≤–Ĺ—Ė —É –Ī—Ė–Ľ—Ć—ą–ĺ—Ā—ā—Ė —ā–ļ–į–Ĺ–ł–Ĺ, —Ė–Ĺ—ą—Ė, –Ĺ–į–Ņ—Ä–ł–ļ–Ľ–į–ī –ļ–į—Ä–Ī–į–ľ–ĺ—ó–Ľ—Ą–ĺ—Ā—Ą–į—ā—Ā–ł–Ĺ—ā–į–∑–į I, –ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ—ā—Ä–į–Ĺ—Ā–ļ–į—Ä–Ī–į–ľ–ĺ—ó–Ľ–į–∑–į —ā–į N-–į—Ü–Ķ—ā–ł–Ľ–≥–Ľ—É—ā–į–ľ–į—ā—Ā–ł–Ĺ—ā–į–∑–į (—Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā, –Ĺ–Ķ–ĺ–Ī—Ö—Ė–ī–Ĺ–ł–Ļ –ī–Ľ—Ź —Ā–ł–Ĺ—ā–Ķ–∑—É N-–į—Ü–Ķ—ā–ł–Ľ–≥–Ľ—É—ā–į–ľ–į—ā—É), –Ķ–ļ—Ā–Ņ—Ä–Ķ—Ā—É—é—ā—Ć—Ā—Ź —ā—Ė–Ľ—Ć–ļ–ł –Ņ–Ķ—á—Ė–Ĺ–ļ–ĺ—é —ā–į —ā–ĺ–Ĺ–ļ–ĺ—é –ļ–ł—ą–ļ–ĺ—é, –Ņ—Ä–ĺ–Ľ—Ė–Ĺ–ĺ–ļ—Ā–ł–ī–į–∑–į ‚ÄĒ –ļ—Ä—Ė–ľ —Ü–ł—Ö –ĺ—Ä–≥–į–Ĺ—Ė–≤ —ā–į–ļ–ĺ–∂ —Ė –Ĺ–ł—Ä–ļ–į–ľ–ł, –į –≥–Ľ—É—ā–į–ľ–į—ā–ļ—Ė–Ĺ–į–∑–į —Ė ő≥-–≥–Ľ—É—ā–į–ľ—Ė–Ľ—Ą–ĺ—Ā—Ą–į—ā—Ä–Ķ–ī—É–ļ—ā–į–∑–į –Ĺ–į—Ź–≤–Ĺ—Ė —ā—Ė–Ľ—Ć–ļ–ł –≤ –Ķ–Ĺ—ā–Ķ—Ä–ĺ—Ü–ł—ā–į—Ö . –Ę–į–ļ–ł–ľ —á–ł–Ĺ–ĺ–ľ –ī–Ķ—Ź–ļ—Ė —Ä–Ķ–į–ļ—Ü—Ė—ó —Ü—Ć–ĺ–≥–ĺ –Ī—Ė–ĺ—Ö—Ė–ľ—Ė—á–Ĺ–ĺ–≥–ĺ —ą–Ľ—Ź—Ö—É –ľ–ĺ–∂—É—ā—Ć –≤—Ė–ī–Ī—É–≤–į—ā–ł—Ā—Ć —ā—Ė–Ľ—Ć–ļ–ł –≤ –ĺ–ļ—Ä–Ķ–ľ–ł—Ö –ĺ—Ä–≥–į–Ĺ–į—Ö. –Ď—Ė–Ľ—Ć—ą—Ė—Ā—ā—Ć –Ī—Ė–ĺ—Ā–ł–Ĺ—ā–Ķ–∑—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≤ –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ—Ė –≤—Ė–ī–Ī—É–≤–į—Ē—ā—Ć—Ā—Ź –∑–į–≤–ī—Ź–ļ–ł –ľ–Ķ—ā–į–Ī–ĺ–Ľ—Ė—á–Ĺ—Ė–Ļ –ļ–ĺ–Ľ–į–Ī–ĺ—Ä–į—Ü—Ė—ó –ľ—Ė–∂ —ā–ĺ–Ĺ–ļ–ĺ—é –ļ–ł—ą–ļ–ĺ—é —ā–į –Ĺ–ł—Ä–ļ–į–ľ–ł, –≤—Ė–ī–ĺ–ľ—Ė–Ļ —ā–į–ļ–ĺ–∂ —Ź–ļ –ļ–ł—ą–ļ–ĺ–≤–ĺ-–Ĺ–ł—Ä–ļ–ĺ–≤–į –≤—Ė—Ā—Ć (–į–Ĺ–≥–Ľ. intestinal-renal axis )[ 5]
–ö–Ľ—Ė—ā–ł–Ĺ–ł —ā–ĺ–Ĺ–ļ–ĺ—ó –ļ–ł—ą–ļ–ł –Ņ–ĺ–≥–Ľ–ł–Ĺ–į—é—ā—Ć –≥–Ľ—É—ā–į–ľ—Ė–Ĺ —Ė –≥–Ľ—É—ā–į–ľ–į—ā, –į —ā–į–ļ–ĺ–∂ –Ņ—Ä–ĺ–Ľ—Ė–Ĺ, —Ė–∑ –ļ—Ä–ĺ–≤—Ė —ā–į –Ņ–ĺ—Ä–ĺ–∂–Ĺ–ł–Ĺ–ł –ļ–ł—ą–ļ–ł, —É —ó—Ö–Ĺ—Ė—Ö –ľ—Ė—ā–ĺ—Ö–ĺ–Ĺ–ī—Ä—Ė—Ź—Ö –∑–Ĺ–į—á–Ĺ–į —á–į—Ā—ā–ł–Ĺ–į —Ü–ł—Ö –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā –Ņ–Ķ—Ä–Ķ—ā–≤–ĺ—Ä—é—Ē—ā—Ć—Ā—Ź —É —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ, –≤ –Ķ–Ĺ—ā–Ķ—Ä–ĺ—Ü–ł—ā–į—Ö —ā–į–ļ–ĺ–∂ –ľ–ĺ–∂–Ķ —Ā–ł–Ĺ—ā–Ķ–∑—É–≤–į—ā–ł—Ā—Ć –ī–Ķ—Ź–ļ–į –ļ—Ė–Ľ—Ć–ļ—Ė—Ā—ā—Ć –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É. –¶–ł—ā—Ä—É–Ľ—Ė–Ĺ –≤–ł–≤—Ė–Ľ—Ć–Ĺ—Ź—Ē—ā—Ć—Ā—Ź —É –ļ—Ä–ĺ–≤, –∑–≤—Ė–ī–ļ–ł –ľ–ĺ–∂–Ķ –Ņ–ĺ–≥–Ľ–ł–Ĺ–į—ā–ł—Ā—Ć —Ė–Ĺ—ą–ł–ľ–ł –ĺ—Ä–≥–į–Ĺ–į–ľ–ł. –Ě–į–Ļ–į–ļ—ā–ł–≤–Ĺ—Ė—ą–Ķ –Ļ–ĺ–≥–ĺ –≤–ł–ļ–ĺ—Ä–ł—Ā—ā–ĺ–≤—É—é—ā—Ć –Ĺ–ł—Ä–ļ–ł, –Ĺ–į —Ź–ļ—Ė –Ņ—Ä–ł–Ņ–į–ī–į—Ē –Ņ—Ä–ł–Ī–Ľ–ł–∑–Ĺ–ĺ 60 % —Ā—É–ľ–į—Ä–Ĺ–ĺ–≥–ĺ —Ā–ł–Ĺ—ā–Ķ–∑—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≤ –ī–ĺ—Ä–ĺ—Ā–Ľ–ł—Ö —Ā—Ā–į–≤—Ü—Ė–≤. –ü–Ķ—Ä–Ķ—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ—É –≤ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ –≤—Ė–ī–Ī—É–≤–į—Ē—ā—Ć—Ā—Ź –∑–į–≤–ī—Ź–ļ–ł –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ė –ź–°–° —ā–į –ź–°–õ. –Ě–į–Ļ–Ī—Ė–Ľ—Ć—ą–į –ļ–ĺ–Ĺ—Ü–Ķ–Ĺ—ā—Ä–į—Ü—Ė—Ź —Ü–ł—Ö —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā—Ė–≤ —Ā–Ņ–ĺ—Ā—ā–Ķ—Ä—Ė–≥–į—Ē—ā—Ć—Ā—Ź —É –Ņ—Ä–ĺ–ļ—Ā–ł–ľ–į–Ľ—Ć–Ĺ–ł—Ö –∑–≤–ł–≤–ł—Ā—ā–ł—Ö –ļ–į–Ĺ–į–Ľ—Ć—Ü—Ź—Ö[ 5]
–ü—Ä–ĺ—Ą—Ė–Ľ—Ć —Ā–ł–Ĺ—ā–Ķ–∑—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É —Ä—Ė–∑–Ĺ–ł–ľ–ł –ĺ—Ä–≥–į–Ĺ–į–ľ–ł –∑–ľ—Ė–Ĺ—é—Ē—ā—Ć—Ā—Ź —Ė–∑ –≤—Ė–ļ–ĺ–ľ: –≤—Ė–ī—Ä–į–∑—É –Ņ—Ė—Ā–Ľ—Ź –Ĺ–į—Ä–ĺ–ī–∂–Ķ–Ĺ–Ĺ—Ź –≤–Ķ—Ā—Ć –Ī—Ė–ĺ—Ö—Ė–ľ—Ė—á–Ĺ–ł–Ļ —ą–Ľ—Ź—Ö —É—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź —Ü—Ė—Ē—ó –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–ł –≤—Ė–ī–Ī—É–≤–į—Ē—ā—Ć—Ā—Ź —É —ā–ĺ–Ĺ–ļ—Ė–Ļ –ļ–ł—ą—Ü—Ė, –Ņ—Ė–∑–Ĺ—Ė—ą–Ķ –≤ –Ķ–Ĺ—ā–Ķ—Ä–ĺ—Ü–ł—ā–į—Ö –∑–Ī—Ė–Ľ—Ć—ą—É—Ē—ā—Ć—Ā—Ź –Ķ–ļ—Ā–Ņ—Ä–Ķ—Ā—Ė—Ź –į—Ä–≥—Ė–Ĺ–į–∑–ł —Ė –ĺ—Ā–Ĺ–ĺ–≤–Ĺ–ł–ľ —ó—Ö–Ĺ—Ė–ľ –Ņ—Ä–ĺ–ī—É–ļ—ā–ĺ–ľ —Ā—ā–į—Ē —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ. –ě–ī–Ĺ–ĺ—á–į—Ā–Ĺ–ĺ –≤ –Ĺ–ł—Ä–ļ–į—Ö –∑—Ä–ĺ—Ā—ā–į—Ē –į–ļ—ā–ł–≤–Ĺ—Ė—Ā—ā—Ć —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā—Ė–≤, –Ĺ–Ķ–ĺ–Ī—Ö—Ė–ī–Ĺ–ł—Ö –ī–Ľ—Ź –Ņ–Ķ—Ä–Ķ—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ—É –≤ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ[ 5]
–ě—Ā–Ĺ–ĺ–≤–Ĺ–ł–ľ —á–ł–Ĺ–Ĺ–ł–ļ–ĺ–ľ, —Č–ĺ –≤–ł–∑–Ĺ–į—á–į—Ē –į–ļ—ā–ł–≤–Ĺ—Ė—Ā—ā—Ć –Ī—Ė–ĺ—Ā–ł–Ĺ—ā–Ķ–∑—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –Ĺ–ł—Ä–ļ–į–ľ–ł, —Ē –ļ–ĺ–Ĺ—Ü–Ķ–Ĺ—ā—Ä–į—Ü—Ė—Ź —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ—É –≤ –ļ—Ä–ĺ–≤—Ė, –ĺ—Ā–ļ—Ė–Ľ—Ć–ļ–ł —ā–į —á–į—Ā—ā–ł–Ĺ–į –Ī—Ė–ĺ—Ö—Ė–ľ—Ė—á–Ĺ–ĺ–≥–ĺ —ą–Ľ—Ź—Ö—É, —Ź–ļ–į –≤—Ė–ī–Ī—É–≤–į—Ē—ā—Ć—Ā—Ź —É —ā–ĺ–Ĺ–ļ—Ė–Ļ –ļ–ł—ą—Ü—Ė, –Ņ—Ä–ĺ—ā—Ė–ļ–į—Ē –∑–Ĺ–į—á–Ĺ–ĺ –Ņ–ĺ–≤—Ė–Ľ—Ć–Ĺ—Ė—ą–Ķ. –ü—Ä–ĺ—ā–Ķ –Ķ–ļ—Ā–Ņ–Ķ—Ä–ł–ľ–Ķ–Ĺ—ā–į–Ľ—Ć–Ĺ—Ė –ī–į–Ĺ—Ė —Ā–≤—Ė–ī—á–į—ā—Ć, —Č–ĺ —É—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ—É –Ĺ–Ķ —Ē —Ā—ā—Ä–ĺ–≥–ĺ —Ä–Ķ–≥—É–Ľ—Ć–ĺ–≤–į–Ĺ–ł–ľ –Ņ—Ä–ĺ—Ü–Ķ—Ā–ĺ–ľ. –ó–ĺ–ļ—Ä–Ķ–ľ–į, –Ļ–ĺ–≥–ĺ —Ė–Ĺ—ā–Ķ–Ĺ—Ā–ł–≤–Ĺ—Ė—Ā—ā—Ć —É —Č—É—Ä—Ė–≤ —ā–į —Ā–≤–ł–Ĺ–Ķ–Ļ –Ĺ–Ķ –∑–Ī—Ė–Ľ—Ć—ą—É—Ē—ā—Ć—Ā—Ź –∑–į —É–ľ–ĺ–≤ –≤–∂–ł–≤–į–Ĺ–Ĺ—Ź –ī—Ė—Ē—ā–ł –Ī—Ė–ī–Ĺ–ĺ—ó –Ĺ–į –į—Ä–≥—Ė–Ĺ—Ė–Ĺ, –Ĺ–į–≤—Ė—ā—Ć –ļ–ĺ–Ľ–ł –Ņ–ĺ–Ņ–Ķ—Ä–Ķ–ī–Ĺ–ł–ļ–ł (–≥–Ľ—É—ā–į–ľ—Ė–Ĺ, –≥–Ľ—É—ā–į–ľ–į—ā) –Ĺ–į—Ź–≤–Ĺ—Ė —É –≤–Ķ–Ľ–ł–ļ—Ė–Ļ –ļ—Ė–Ľ—Ć–ļ–ĺ—Ā—ā—Ė. –ü—Ä–ł–Ņ—É—Ā–ļ–į—Ē—ā—Ć—Ā—Ź, —Č–ĺ –ī–Ľ—Ź —Ä–Ķ–≥—É–Ľ—Ź—Ü—Ė—ó –∑–į–≥–į–Ľ—Ć–Ĺ–ĺ–≥–ĺ –≤–ľ—Ė—Ā—ā—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≤ –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ—Ė –Ī—Ė–Ľ—Ć—ą–Ķ –∑–Ĺ–į—á–Ķ–Ĺ–Ĺ—Ź –ľ–į—Ē –ļ–ĺ–Ĺ—ā—Ä–ĺ–Ľ—Ć –Ĺ–į–ī –Ļ–ĺ–≥–ĺ –ļ–į—ā–į–Ī–ĺ–Ľ—Ė–∑–ľ–ĺ–ľ , –Ĺ—Ė–∂ –Ĺ–į–ī –Ī—Ė–ĺ—Ā–ł–Ĺ—ā–Ķ–∑–ĺ–ľ[ 5]
–£ –≥–Ķ–Ņ–į—ā–ĺ—Ü–ł—ā–į—Ö –≤–Ķ–Ľ–ł–ļ–į –ļ—Ė–Ľ—Ć–ļ—Ė—Ā—ā—Ć –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –Ņ–ĺ—Ā—ā—Ė–Ļ–Ĺ–ĺ —É—ā–≤–ĺ—Ä—é—Ē—ā—Ć—Ā—Ź –≤ —Ü–ł–ļ–Ľ—Ė —Ā–Ķ—á–ĺ–≤–ł–Ĺ–ł, –Ņ—Ä–ĺ—ā–Ķ, –≤—Ė–Ĺ –ĺ–ī—Ä–į–∑—É –∂ —Ä–ĺ–∑—Č–Ķ–Ņ–Ľ—é—Ē—ā—Ć—Ā—Ź –į—Ä–≥—Ė–Ĺ–į–∑–ĺ—é. –Ę–į–ļ–ł–ľ —á–ł–Ĺ–ĺ–ľ –ī–Ľ—Ź —ā–ĺ–≥–ĺ, —Č–ĺ–Ī —É –Ņ–Ķ—á—Ė–Ĺ—Ü—Ė –≤—Ė–ī–Ī—É–≤–į–≤—Ā—Ź —Ā—É–ľ–į—Ä–Ĺ–ł–Ļ —Ā–ł–Ĺ—ā–Ķ–∑ —Ü—Ė—Ē—ó –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–ł, —Ü–ł–ļ–Ľ —Ā–Ķ—á–ĺ–≤–ł–Ĺ–ł –Ņ–ĺ–≤–ł–Ĺ–Ķ–Ĺ –Ņ–ĺ–Ņ–ĺ–≤–Ĺ—é–≤–į—ā–ł—Ā—Ć –∑–į —Ä–į—Ö—É–Ĺ–ĺ–ļ —Ė–Ĺ—ą–ł—Ö –ľ–Ķ—ā–į–Ī–ĺ–Ľ—Ė—ā—Ė–≤, —ā–į–ļ–ł—Ö —Ź–ļ –ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ. –ö—Ä—Ė–ľ —ā–ĺ–≥–ĺ, –≤—Ā—Ė —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā–ł —Ü–ł–ļ–Ľ—É —Ā–Ķ—á–ĺ–≤–ł–Ĺ–ł –ĺ—Ä–≥–į–Ĺ—Ė–∑–ĺ–≤–į–Ĺ—Ė —É –ľ–Ķ—ā–į–Ī–ĺ–Ľ–ĺ–Ĺ , –≤ —Ź–ļ–ĺ–ľ—É –Ņ—Ä–ĺ–ľ—Ė–∂–Ĺ—Ė –ľ–Ķ—ā–į–Ī–ĺ–Ľ—Ė—ā–ł –Ĺ–Ķ –≤–ł–≤—Ė–Ľ—Ć–Ĺ—Ź—é—ā—Ć—Ā—Ź —É —Ü–ł—ā–ĺ–Ņ–Ľ–į–∑–ľ—É (—á–ł –ľ–į—ā—Ä–ł–ļ—Ā –ľ—Ė—ā–ĺ—Ö–ĺ–Ĺ–ī—Ä—Ė–Ļ), –į –Ņ–Ķ—Ä–Ķ–ī–į—é—ā—Ć—Ā—Ź –≤—Ė–ī –ĺ–ī–Ĺ–ĺ–≥–ĺ –į–ļ—ā–ł–≤–Ĺ–ĺ–≥–ĺ —Ü–Ķ–Ĺ—ā—Ä—É –ī–ĺ —Ė–Ĺ—ą–ĺ–≥–ĺ. –ü—Ä–ĺ —ā–Ķ, —Č–ĺ –Ņ–Ķ—á—Ė–Ĺ–ļ–į —Ä–ĺ–Ī–ł—ā—Ć –ī—É–∂–Ķ –ľ–į–Ľ–ł–Ļ –≤–ļ–Ľ–į–ī —É —Ā—É–ľ–į—Ä–Ĺ–ł–Ļ —Ā–ł–Ĺ—ā–Ķ–∑ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É (–į–Ī–ĺ –≤–∑–į–≥–į–Ľ—Ė –Ĺ–Ķ —Ä–ĺ–Ī–ł—ā—Ć), —Ā–≤—Ė–ī—á–į—ā—Ć –≤–ł–Ņ–į–ī–ļ–ł —ā—Ä–į–Ĺ—Ā–Ņ–Ľ–į–Ĺ—ā–į—Ü—Ė—ó —Ü—Ć–ĺ–≥–ĺ –ĺ—Ä–≥–į–Ĺ–į –Ľ—é–ī—Ź–ľ —Ė–∑ –≤—Ä–ĺ–ī–∂–Ķ–Ĺ–ł–ľ–ł –ī–Ķ—Ą–Ķ–ļ—ā–į–ľ–ł —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā—Ė–≤ —Ü–ł–ļ–Ľ—É —Ā–Ķ—á–ĺ–≤–ł–Ĺ–ł. –Ě–į–≤—Ė—ā—Ć –Ņ—Ė—Ā–Ľ—Ź –ĺ—ā—Ä–ł–ľ–į–Ĺ–Ĺ—Ź –∑–ī–ĺ—Ä–ĺ–≤–ĺ—ó –Ņ–Ķ—á—Ė–Ĺ–ļ–ł –≤–ĺ–Ĺ–ł –Ņ–ĺ–≤–ł–Ĺ–Ĺ—Ė –ĺ—ā—Ä–ł–ľ—É–≤–į—ā–ł –≤–Ķ—Ā—Ć –į—Ä–≥—Ė–Ĺ—Ė–Ĺ —Ė–∑ –ī—Ė—Ē—ā–ł[ 5]
–ź—Ä–≥—Ė–Ĺ—Ė–Ĺ —Ē –ĺ—Ā–Ĺ–ĺ–≤–Ĺ–ł–ľ –ī–∂–Ķ—Ä–Ķ–Ľ–ĺ–ľ –ī–Ľ—Ź —Ā–ł–Ĺ—ā–Ķ–∑—É NO —É –Ī–į–≥–į—ā—Ć–ĺ—Ö —ā–ł–Ņ–į—Ö –ļ–Ľ—Ė—ā–ł–Ĺ, —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā NO-—Ā–ł–Ĺ—ā–į–∑–į (iNOS ) –Ņ–Ķ—Ä–Ķ—ā–≤–ĺ—Ä—é—Ē –Ļ–ĺ–≥–ĺ –Ĺ–į –ĺ–ļ—Ā–ł–ī –Ě—Ė—ā—Ä–ĺ–≥–Ķ–Ĺ—É (II) —ā–į —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ. –ě—Ā—ā–į–Ĺ–Ĺ—Ė–Ļ –ľ–ĺ–∂–Ķ –∑–Ĺ–ĺ–≤—É –Ņ–Ķ—Ä–Ķ—ā–≤–ĺ—Ä—é–≤–į—ā–ł—Ā—Ć –≤ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ, —É —ą–Ľ—Ź—Ö—É –≤—Ė–ī–ĺ–ľ–ĺ–ľ—É —Ź–ļ —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ/NO —Ü–ł–ļ–Ľ (–į–Ī–ĺ —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ/–į—Ä–≥—Ė–Ĺ—Ė–Ĺ-—Ü–ł–ļ–Ľ), –ĺ—Ā–Ĺ–ĺ–≤–Ĺ–ł–ľ–ł —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā–į–ľ–ł —Ź–ļ–ĺ–≥–ĺ —Ē –ź–°–° —ā–į –ź–°–õ. –ü—Ä–ĺ—ā–Ķ —Ė–Ĺ—ā–Ķ–Ĺ—Ā–ł–≤–Ĺ—Ė—Ā—ā—Ć —Ü—Ć–ĺ–≥–ĺ —ą–Ľ—Ź—Ö—É –∑–Ĺ–į—á–Ĺ–ĺ –Ĺ–ł–∂—á–į, –Ĺ—Ė–∂ —Ü–ł–ļ–Ľ—É —Ā–Ķ—á–ĺ–≤–ł–Ĺ–ł —É –Ņ–Ķ—á—Ė–Ĺ—Ü—Ė, –≤–Ĺ–į—Ā–Ľ—Ė–ī–ĺ–ļ —á–ĺ–≥–ĺ –≤ –ļ–Ľ—Ė—ā–ł–Ĺ–į—Ö –ľ–ĺ–∂–Ķ –Ĺ–į–ļ–ĺ–Ņ–ł—á—É–≤–į—ā–ł—Ā—Ć —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ. –ü—Ä–ł—á–ł–Ĺ–ĺ—é —Ü—Ć–ĺ–≥–ĺ –ľ–ĺ–∂–Ķ –Ī—É—ā–ł –∑–Ĺ–ł–∂–Ķ–Ĺ–į –į–ļ—ā–ł–≤–Ĺ—Ė—Ā—ā—Ć –ź–°–° —É –Ņ–ĺ—Ä—Ė–≤–Ĺ—Ź–Ĺ–Ĺ—Ė —Ė–∑ iNOS –į–Ī–ĺ/—Ė –Ī—Ä–į–ļ–ĺ–ľ –ļ–į–Ĺ–į–Ľ—é–≤–į–Ĺ–Ĺ—Ź —Ā—É–Ī—Ā—ā—Ä–į—ā—Ė–≤ –ľ—Ė–∂ —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā–į–ľ–ł[ 5]
–ě—Ā–Ĺ–ĺ–≤–Ĺ–ł–Ļ —ą–Ľ—Ź—Ö –Ī—Ė–ĺ—Ā–ł–Ĺ—ā–Ķ–∑—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≤ –Ī–į–ļ—ā–Ķ—Ä—Ė–Ļ –£ –Ī–į–ļ—ā–Ķ—Ä—Ė–Ļ —ą–Ľ—Ź—Ö –Ī—Ė–ĺ—Ā–ł–Ĺ—ā–Ķ–∑—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É —Ė–∑ –≥–Ľ—É—ā–į–ľ–į—ā—É de novo —Ā—Ö–ĺ–∂–ł–Ļ –ī–ĺ —ā–į–ļ–ĺ–≥–ĺ —É —Ā—Ā–į–≤—Ü—Ė–≤, –Ņ—Ä–ĺ—ā–Ķ –≤—Ė–Ĺ –ľ–ĺ–∂–Ķ –Ņ—Ä–ĺ—ā—Ė–ļ–į—ā–ł –Ī—Ė–Ľ—Ć—ą –Ķ—Ą–Ķ–ļ—ā–ł–≤–Ĺ–ĺ, —á–Ķ—Ä–Ķ–∑ —ā–Ķ, —Č–ĺ –į–ľ—Ė–Ĺ–ĺ–≥—Ä—É–Ņ–į –≤ őĪ-–Ņ–ĺ–Ľ–ĺ–∂–Ķ–Ĺ–Ĺ—Ė –≥–Ľ—É—ā–į–ľ–į—ā—É –∑–į—Ö–ł—Č–į—Ē—ā—Ć—Ā—Ź –į—Ü–Ķ—ā–ł–Ľ—Ć–Ĺ–ĺ—é –≥—Ä—É–Ņ–ĺ—é, —á–Ķ—Ä–Ķ–∑ —Č–ĺ —Ü–ł–ļ–Ľ—Ė–∑–į—Ü—Ė—Ź —Ā–Ķ–ľ—Ė–į–Ľ—Ć–ī–Ķ–≥—Ė–ī—É —Ā—ā–į—Ē –Ĺ–Ķ–ľ–ĺ–∂–Ľ–ł–≤–ĺ—é. –ü–Ķ—Ä—ą–ĺ—é —Ä–Ķ–į–ļ—Ü—Ė—Ē—é —Ü—Ć–ĺ–≥–ĺ —ą–Ľ—Ź—Ö—É —Ē —É—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź N-–į—Ü–Ķ—ā–ł–Ľ–≥–Ľ—É—ā–į–ľ–į—ā—É , –≤–Ĺ–į—Ā–Ľ—Ė–ī–ĺ–ļ –Ņ–Ķ—Ä–Ķ–Ĺ–Ķ—Ā–Ķ–Ĺ–Ĺ—Ź –į—Ü–Ķ—ā–ł–Ľ—Ć–Ĺ–ĺ—ó –≥—Ä—É–Ņ–ł —Ė–∑ –į—Ü–Ķ—ā–ł–Ľ-–ö–ĺ–ź —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā–ĺ–ľ N-–į—Ü–Ķ—ā–ł–Ľ–≥–Ľ—É—ā–į–ľ–į—ā—Ā–ł–Ĺ—ā–į–∑–ĺ—é. –Ē–į–Ľ—Ė N-–į—Ü–Ķ—ā–ł–Ľ–≥–Ľ—É—ā–į–ľ–į—ā –į–ļ—ā–ł–≤—É—Ē—ā—Ć—Ā—Ź –Ņ—Ä–ł—Ē–ī–Ĺ–į–Ĺ–Ĺ—Ź–ľ —Ą–ĺ—Ā—Ą–į—ā–Ĺ–ĺ—ó –≥—Ä—É–Ņ–ł —Ė –≤—Ė–ī–Ĺ–ĺ–≤–Ľ—é—Ē—ā—Ć—Ā—Ź –ī–ĺ N-–į—Ü–Ķ—ā–ł–Ľ–≥–Ľ—É—ā–į–ľ–į—ā-ő≥-—Ā–Ķ–ľ—Ė–į–Ľ—Ć–ī–Ķ–≥—Ė–ī—É. –ě—Ā—ā–į–Ĺ–Ĺ—Ė–Ļ –≤—Ā—ā—É–Ņ–į—Ē –≤ —Ä–Ķ–į–ļ—Ü—Ė—é –Ņ–Ķ—Ä–Ķ–į–ľ—Ė–Ĺ—É–≤–į–Ĺ–Ĺ—Ź —Ė–∑ –≥–Ľ—É—ā–į–ľ–į—ā–ĺ–ľ, –ĺ–ī–Ĺ–ł–ľ —Ė–∑ –Ņ—Ä–ĺ–ī—É–ļ—ā—Ė–≤ —Ź–ļ–ĺ—ó —Ē N-–į—Ü–Ķ—ā–ł–Ľ–ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ , —Ź–ļ–ł–Ļ –Ņ–Ķ—Ä–Ķ—ā–≤–ĺ—Ä—é—Ē—ā—Ć—Ā—Ź –≤ –ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ —ą–Ľ—Ź—Ö–ĺ–ľ –≤—Ė–ī—Č–Ķ–Ņ–Ľ–Ķ–Ĺ–Ĺ—Ź –į—Ü–Ķ—ā–ł–Ľ—Ć–Ĺ–ĺ—ó –≥—Ä—É–Ņ–ł (—Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā N-–į—Ü–Ķ—ā–ł–Ľ–ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ–į–∑–į ). –ü–ĺ–ī–į–Ľ—Ć—ą—Ė –Ņ–Ķ—Ä–Ķ—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź –ĺ—Ä—Ė–Ĺ—ā–ł–Ĺ—É –į–Ĺ–į–Ľ–ĺ–≥—Ė—á–Ĺ—Ė –ī–ĺ —ā–ł—Ö, —Č–ĺ –≤—Ė–ī–Ī—É–≤–į—é—ā—Ć—Ā—Ź —É —Ā—Ā–į–≤—Ü—Ė–≤.
–ź—Ä–≥—Ė–Ĺ—Ė–Ĺ –Ņ–ĺ–≥–Ľ–ł–Ĺ–į—Ē—ā—Ć—Ā—Ź –ļ–Ľ—Ė—ā–ł–Ĺ–į–ľ–ł —Ė–∑ –Ņ–Ľ–į–∑–ľ–ł, –Ļ–ĺ–≥–ĺ —á–į—Ā –Ņ—Ė–≤–∂–ł—ā—ā—Ź —É –ļ—Ä–ĺ–≤—Ė –ī–ĺ—Ä–ĺ—Ā–Ľ–ĺ—ó —Ā–≤–ł–Ĺ—Ė —Ā—ā–į–Ĺ–ĺ–≤–ł—ā—Ć 1,06 –≥–ĺ–ī, –≤–į–≥—Ė—ā–Ĺ–ĺ—ó ‚ÄĒ 0,75 –≥–ĺ–ī, –į –Ĺ–Ķ–ĺ–Ĺ–į—ā–į–Ľ—Ć–Ĺ–ĺ—ó ‚ÄĒ 0,65 –≥–ĺ–ī[ 3] –ľ–Ķ–ľ–Ī—Ä–į–Ĺ—É –∑–į–Ī–Ķ–∑–Ņ–Ķ—á—É—Ē—ā—Ć—Ā—Ź –ĺ–ī–Ĺ—Ė—Ē—é —Ė–∑ —Ā–ł—Ā—ā–Ķ–ľ y+ , bo,+ , Bo,+ , y+ L. –ó —Ź–ļ–ł—Ö –Ĺ–į–Ļ–≤–į–∂–Ľ–ł–≤—Ė—ą–ĺ—é —Ē –Ņ–Ķ—Ä—ą–į ‚ÄĒ y+ ‚ÄĒ –≤–ł—Ā–ĺ–ļ–ĺ–į—Ą—Ė–Ĺ–Ĺ–ł–Ļ Na -–Ĺ–Ķ–∑–į–Ľ–Ķ–∂–Ĺ–ł–Ļ —ā—Ä–į–Ĺ—Ā–Ņ–ĺ—Ä—ā–Ķ—Ä –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É, –Ľ—Ė–∑–ł–Ĺ—É —ā–į –ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ—É. –ü–Ķ—Ä–Ķ–Ľ—Ė—á–Ķ–Ĺ—Ė —Ā–ł—Ā—ā–Ķ–ľ–ł —ā—Ä–į–Ĺ—Ā–Ņ–ĺ—Ä—ā—É –Ķ–ļ—Ā–Ņ—Ä–Ķ—Ā—É—é—ā—Ć—Ā—Ź —ā–ļ–į–Ĺ–ł–Ĺ–ĺ—Ā–Ņ–Ķ—Ü–ł—Ą—Ė—á–Ĺ–ĺ, –Ĺ–į–Ņ—Ä–ł–ļ–Ľ–į–ī y+ –Ĺ–į—Ź–≤–Ĺ–į –ľ–į–Ļ–∂–Ķ —É –≤—Ā—Ė—Ö –ļ–Ľ—Ė—ā–ł–Ĺ–į—Ö –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ—É, –ļ—Ä—Ė–ľ –≥–Ķ–Ņ–į—ā–ĺ—Ü–ł—ā—Ė–≤. –ö—Ė–Ľ—Ć–ļ—Ė—Ā—ā—Ć —ā—Ä–į–Ĺ—Ā–Ņ–ĺ—Ä—ā–Ķ—Ä—Ė–≤ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –Ņ—Ė–ī–Ľ—Ź–≥–į—Ē —Ä–Ķ–≥—É–Ľ—é–≤–į–Ĺ–Ĺ—é, –∑–ĺ–ļ—Ä–Ķ–ľ–į –≤–ĺ–Ĺ–į –ľ–ĺ–∂–Ķ –Ī—É—ā–ł –∑–Ī—Ė–Ľ—Ć—ą–Ķ–Ĺ–į —É –≤—Ė–ī–Ņ–ĺ–≤—Ė–ī—Ć –Ĺ–į –ī—Ė—é –Ī–į–ļ—ā–Ķ—Ä—Ė–Ļ–Ĺ–ł—Ö –Ķ–Ĺ–ī–ĺ—ā–ĺ–ļ—Ā–ł–Ĺ—Ė–≤ –į–Ī–ĺ —Ü–ł—ā–ĺ–ļ—Ė–Ĺ—Ė–≤ –∑–į–Ņ–į–Ľ–Ķ–Ĺ–Ĺ—Ź [ 5]
–ź—Ä–≥—Ė–Ĺ—Ė–Ĺ –ļ–ĺ–ī—É—Ē—ā—Ć—Ā—Ź —ą—Ė—Ā—ā—Ć–ľ–į –ļ–ĺ–ī–ĺ–Ĺ–į–ľ–ł –ľ–†–Ě–ö: –¶–ď–ź, –¶–ď–£, –¶–ď–ď, –¶–ď–¶, –ź–ď–ź —ā–į –ź–ď–ď. –Į–ļ—Ė –≤—Ā—Ė —Ė–Ĺ—ą—Ė –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–ł, –Ņ–Ķ—Ä–Ķ–ī –≤–ļ–Ľ—é—á–Ķ–Ĺ–Ĺ—Ź–ľ —É –Ī—Ė–Ľ–ĺ–ļ –≤—Ė–Ĺ –Ī—É—ā–ł –į–ļ—ā–ł–≤–ĺ–≤–į–Ĺ–ł–ľ —Ė –Ņ—Ä–ł—Ē–ī–Ĺ–į–Ĺ–ł–ľ –ī–ĺ –≤—Ė–ī–Ņ–ĺ–≤—Ė–ī–Ĺ–ĺ—ó —ā–†–Ě–ö . –¶—Ė —Ä–Ķ–į–ļ—Ü—Ė—ó –∑–į–Ī–Ķ–∑–Ņ–Ķ—á—É—Ē —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā –į—Ä–≥—Ė–Ĺ—Ė–Ľ-—ā–†–Ě–ö-—Ā–ł–Ĺ—ā–į–∑–į . –°–Ņ–Ķ—Ä—ą—É –≤—Ė–ī–Ī—É–≤–į—Ē—ā—Ć—Ā—Ź –į–ī–Ķ–Ĺ—Ė–Ľ—é–≤–į–Ĺ–Ĺ—Ź –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –Ņ–ĺ –ļ–į—Ä–Ī–ĺ–ļ—Ā–ł–Ľ—Ć–Ĺ—Ė–Ļ –≥—Ä—É–Ņ—Ė, –Ņ—Ė–∑–Ĺ—Ė—ą–Ķ –į–ī–Ķ–Ĺ—Ė–Ľ –∑–į–ľ—Ė—Č—É—Ē—ā—Ć—Ā—Ź –Ĺ–į –≤—Ė–ī–Ņ–ĺ–≤—Ė–ī–Ĺ—É —ā–†–Ě–ö. –£—ā–≤–ĺ—Ä–Ķ–Ĺ–į —Ā–Ņ–ĺ–Ľ—É–ļ–į –į—Ä–≥—Ė–Ĺ—Ė–Ľ-—ā–†–Ě–ö –ľ–ĺ–∂–Ķ –Ī—É—ā–ł –≤–ł–ļ–ĺ—Ä–ł—Ā—ā–į–Ĺ–į —É –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ė —ā—Ä–į–Ĺ—Ā–Ľ—Ź—Ü—Ė—ó , —Č–ĺ –≤—Ė–ī–Ī—É–≤–į—Ē—ā—Ć—Ā—Ź –Ĺ–į —Ä–ł–Ī–ĺ—Ā–ĺ–ľ–į—Ö .
–ź–Ĺ–į–Ľ—Ė–∑ –Ī—Ė–Ľ—Ć—ą–Ķ 1150 –Ī—Ė–Ľ–ļ—Ė–≤ –Ņ–ĺ–ļ–į–∑–į–≤, —Č–ĺ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ —Ā—ā–į–Ĺ–ĺ–≤–ł—ā—Ć –Ī–Ľ–ł–∑—Ć–ļ–ĺ 5,1 % —É—Ā—Ė—Ö –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā –≤ —ó—Ö —Ā–ļ–Ľ–į–ī—Ė. –ü—Ė–ī–≤–ł—Č–Ķ–Ĺ–ł–Ļ –Ļ–ĺ–≥–ĺ –≤–ľ—Ė—Ā—ā —Ā–Ņ–ĺ—Ā—ā–Ķ—Ä—Ė–≥–į—Ē—ā—Ć—Ā—Ź —É —ā–ł—Ö –Ī—Ė–Ľ–ļ–į—Ö, —Č–ĺ –≤–∑–į—Ē–ľ–ĺ–ī—Ė—é—ā—Ć —Ė–∑ –Ĺ–Ķ–≥–į—ā–ł–≤–Ĺ–ĺ –∑–į—Ä—Ź–ī–∂–Ķ–Ĺ–ł–ľ–ł –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–į–ľ–ł, —ā–į–ļ–ł–ľ–ł —Ź–ļ –Ē–Ě–ö, –∑–ĺ–ļ—Ä–Ķ–ľ–į –≤ –Ņ—Ä–ĺ—ā–į–ľ—Ė–Ĺ —Ā–Ņ–Ķ—Ä–ľ–ł –Ľ–ĺ—Ā–ĺ—Ā—Ź (—Ā–į–Ľ—Ć–ľ—Ė–Ĺ) –ľ—Ė—Ā—ā–ł—ā—Ć 85,2 % —Ü—Ė—Ē—ó –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–ł. –ě—Ā–Ĺ–ĺ–≤–Ĺ—Ė –≤–Ľ–į—Ā—ā–ł–≤–ĺ—Ā—ā—Ė –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É —ā–į–ļ–ĺ–∂ —Ā–Ņ—Ä–ł—Ź—é—ā—Ć —Ą–ĺ—Ä–ľ—É–≤–į–Ĺ–Ĺ—é –Ņ—Ä–ĺ—Ā—ā–ĺ—Ä–ĺ–≤–ĺ—ó —Ā—ā—Ä—É–ļ—ā—É—Ä–ł –Ņ–ĺ–Ľ—Ė–Ņ–Ķ–Ņ—ā–ł–ī—Ė–≤, –∑–į–≤–ī—Ź–ļ–ł —É—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—é —Ė–ĺ–Ĺ–Ĺ–ł—Ö –∑–≤'—Ź–∑–ļ—Ė–≤ —Ė–∑ –≤—Ė–Ľ—Ć–Ĺ–ł–ľ–ł –ļ–į—Ä–Ī–ĺ–ļ—Ā–ł–Ľ—Ć–Ĺ–ł–ľ–ł –≥—Ä—É–Ņ–į–ľ–ł —Ė–Ĺ—ą–ł—Ö –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā.
–ü—Ė—Ā–Ľ—Ź –≤–ļ–Ľ—é—á–Ķ–Ĺ–Ĺ—Ź –ī–ĺ —Ā–ļ–Ľ–į–ī—É –Ņ–ĺ–Ľ—Ė–Ņ–Ķ–Ņ—ā–ł–ī—É –∑–į–Ľ–ł—ą–ĺ–ļ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –ľ–ĺ–∂–Ķ –∑–į–∑–Ĺ–į–≤–į—ā–ł –ľ–ĺ–ī–ł—Ą—Ė–ļ–į—Ü—Ė—ó, –∑–ĺ–ļ—Ä–Ķ–ľ–į –Ņ–Ķ–Ņ—ā–ł–ī–ł–Ľ–į—Ä–≥—Ė–Ĺ—Ė–Ĺ–ī–Ķ–į–ľ—Ė–Ĺ–į–∑–ł (PAD) –∑–ī–į—ā–Ĺ—Ė –Ņ–Ķ—Ä–Ķ—ā–≤–ĺ—Ä—é–≤–į—ā–ł –Ļ–ĺ–≥–ĺ —É –∑–į–Ľ–ł—ą–ĺ–ļ —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ—É[ 16]
–ě–ļ—Ä—Ė–ľ —ā–ĺ–≥–ĺ, —Č–ĺ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ —Ē –ĺ–ī–Ĺ—Ė—Ē—é —Ė–∑ –ī–≤–į–ī—Ü—Ź—ā–ł —Ā—ā–į–Ĺ–ī–į—Ä—ā–Ĺ–ł—Ö –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā, —Č–ĺ –ļ–ĺ–ī—É—é—ā—Ć—Ā—Ź –≥–Ķ–Ĺ–Ķ—ā–ł—á–Ĺ–ł–ľ –ļ–ĺ–ī–ĺ–ľ —Ė –≤—Ö–ĺ–ī—Ź—ā—Ć –ī–ĺ —Ā–ļ–Ľ–į–ī—É –Ī—Ė–Ľ–ļ—Ė–≤, —Ü—Ź –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–į –ľ–į—Ē —Ä—Ź–ī —Ė–Ĺ—ą–ł—Ö —Ą—É–Ĺ–ļ—Ü—Ė–Ļ –≤ –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ—Ė. –ó–ĺ–ļ—Ä–Ķ–ľ–į, –≤–ĺ–Ĺ–į –Ĺ–Ķ–ĺ–Ī—Ö—Ė–ī–Ĺ–į –ī–Ľ—Ź —Ā–ł–Ĺ—ā–Ķ–∑—É —Ā–Ķ—á–ĺ–≤–ł–Ĺ–ł –≤ —É—Ä–Ķ–į—ā–Ķ–Ľ—Ė—á–Ĺ–ł—Ö –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ—Ė–≤, –Ņ—Ä–ĺ–ī—É–ļ—É–≤–į–Ĺ–Ĺ—Ź –ĺ–ļ—Ā–ł–ī—É –Ě—Ė—ā—Ä–ĺ–≥–Ķ–Ĺ—É (II), –Ņ–ĺ–Ľ—Ė–į–ľ—Ė–Ĺ—Ė–≤, –į–≥–ľ–į—ā–ł–Ĺ—É, –ļ—Ä–Ķ–į—ā–ł–Ĺ—É, —ā–į–ļ–ĺ–∂ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ –ľ–ĺ–∂–Ķ –≤–ł—Ā—ā—É–Ņ–į—ā–ł –Ņ–ĺ–Ņ–Ķ—Ä–Ķ–ī–Ĺ–ł–ļ–ĺ–ľ –ī–Ľ—Ź —É—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź —Ė–Ĺ—ą–ł—Ö –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā, –∑–ĺ–ļ—Ä–Ķ–ľ–į –Ņ—Ä–ĺ–Ľ—Ė–Ĺ—É —ā–į –≥–Ľ—É—ā–į–ľ–į—ā—É —Ė –≥–Ľ—É—ā–į–ľ—Ė–Ĺ—É[ 5] [ 4] [ 3]
–Ě–į–Ļ–Ī—Ė–Ľ—Ć—ą–į –ļ—Ė–Ľ—Ć–ļ—Ė—Ā—ā—Ć –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≤ –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ—Ė —Ā—Ā–į–≤—Ü—Ė–≤ –ļ–į—ā–į–Ī–ĺ–Ľ—Ė–∑—É—Ē—ā—Ć—Ā—Ź –∑–į —É—á–į—Ā—ā—Ė —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā—É –į—Ä–≥—Ė–Ĺ–į–∑–ł (–ö–§ 3.5.3.1) –ī–ĺ –ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ—É —ā–į —Ā–Ķ—á–ĺ–≤–ł–Ĺ–ł. –¶—Ź —Ä–Ķ–į–ļ—Ü—Ė—Ź —Ē –ĺ–ī–Ĺ–ł–ľ —Ė–∑ –Ķ—ā–į–Ņ—Ė–≤ —Ü–ł–ļ–Ľ—É —Ā–Ķ—á–ĺ–≤–ł–Ĺ–ł, —Č–ĺ –∑–į–Ī–Ķ–∑–Ņ–Ķ—á—É—Ē –ī–Ķ—ā–ĺ–ļ—Ā–ł—Ą—Ė–ļ–į—Ü—Ė—é –į–ľ—Ė–į–ļ—É –≤ —É—Ä–Ķ–į—ā–Ķ–Ľ—Ė—á–Ĺ–ł—Ö —ā–≤–į—Ä–ł–Ĺ. –¶–ł–ļ–Ľ —Ā–Ķ—á–ĺ–≤–ł–Ĺ–ł –Ņ—Ä–ĺ—ā—Ė–ļ–į—Ē –≥–ĺ–Ľ–ĺ–≤–Ĺ–ł–ľ —á–ł–Ĺ–ĺ–ľ —É –Ņ–Ķ—á—Ė–Ĺ—Ü—Ė, –ī–Ķ —É –≤–Ķ–Ľ–ł–ļ—Ė–Ļ –ļ—Ė–Ľ—Ć–ļ–ĺ—Ā—ā—Ė –Ķ–ļ—Ā–Ņ—Ä–Ķ—Ā—É—Ē—ā—Ć—Ā—Ź –į—Ä–≥—Ė–Ĺ–į–∑–į I (–ĺ–ī–ł–Ĺ —Ė–∑ –ī–≤–ĺ—Ö —Ė–∑–ĺ—Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā—Ė–≤ —É —Ā—Ā–į–≤—Ü—Ė–≤). –Ę–į–ļ–ĺ–∂ –į—Ä–≥—Ė–Ĺ–į–∑–į I –Ĺ–į—Ź–≤–Ĺ–į —É –ļ–Ľ—Ė—ā–ł–Ĺ–į—Ö –Ķ–Ĺ–ī–ĺ—ā–Ķ–Ľ—Ė—é , –Ķ–Ņ—Ė—ā–Ķ–Ľ—Ė–į–Ľ—Ć–Ĺ–ł—Ö –ļ–Ľ—Ė—ā–ł–Ĺ–į—Ö –ľ–ĺ–Ľ–ĺ—á–Ĺ–ł—Ö –∑–į–Ľ–ĺ–∑, –ľ–į–ļ—Ä–ĺ—Ą–į–≥–į—Ö , –į –≤ –Ņ—Ä–ł–ľ–į—ā—Ė–≤ —Č–Ķ —Ė –≤ –Ķ—Ä–ł—ā—Ä–ĺ—Ü–ł—ā–į—Ö . –ź—Ä–≥—Ė–Ĺ–į–∑–į II –Ĺ–į—ā–ĺ–ľ—Ė—Ā—ā—Ć –Ķ–ļ—Ā–Ņ—Ä–Ķ—Ā—É—Ē—ā—Ć—Ā—Ź –ľ–į–Ļ–∂–Ķ —É –≤—Ā—Ė—Ö –ļ–Ľ—Ė—ā–ł–Ĺ–į—Ö, —Č–ĺ –ľ—Ė—Ā—ā—Ź—ā—Ć –ľ—Ė—ā–ĺ—Ö–ĺ–Ĺ–ī—Ä—Ė—ó, –∑–į –≤–ł–Ĺ—Ź—ā–ļ–ĺ–ľ –≥–Ķ–Ņ–į—ā–ĺ—Ü–ł—ā—Ė–≤. –á—ó —Ä–ĺ–Ľ—Ć, –Ļ–ľ–ĺ–≤—Ė—Ä–Ĺ–ĺ, –Ņ–ĺ–Ľ—Ź–≥–į—Ē —É —Ä–Ķ–≥—É–Ľ—Ź—Ü—Ė—ó –ī–ĺ—Ā—ā—É–Ņ—É –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –ī–Ľ—Ź —Ė–Ĺ—ą–ł—Ö —ą–Ľ—Ź—Ö—Ė–≤ –Ļ–ĺ–≥–ĺ –ļ–į—ā–į–Ī–ĺ–Ľ—Ė–∑–ľ—É, –∑–ĺ–ļ—Ä–Ķ–ľ–į –≤–ĺ–Ĺ–į –ľ–ĺ–∂–Ķ –ĺ–Ī–ľ–Ķ–∂—É–≤–į—ā–ł –ī–ĺ—Ā—ā—É–Ņ–Ĺ—Ė—Ā—ā—Ć —Ā—É–Ī—Ā—ā—Ä–į—ā—É –ī–Ľ—Ź —Ā–ł–Ĺ—ā–Ķ–∑—É NO, –∑—Ė–Ī–Ľ—Ć—ą—É–≤–į—ā–ł –ļ—Ė–Ľ—Ć–ļ—Ė—Ā—ā—Ć –ĺ—Ä—Ė–Ĺ—ā–ł–Ĺ—É, –Ĺ–Ķ–ĺ–Ī—Ö—Ė–ī–Ĺ–ĺ–≥–ĺ –ī–Ľ—Ź —É—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź –Ņ–ĺ–Ľ—Ė–į–ľ—Ė–Ĺ—Ė–≤ (–Ņ—É—ā—Ä–Ķ—Ā—Ü–ł–Ĺ—É , —Ā–Ņ–Ķ—Ä–ľ—Ė–ī–ł–Ĺ—É —ā–į —Ā–Ņ–Ķ—Ä–ľ—Ė–Ĺ—É ) —ā–į –Ņ—Ä–ĺ–Ľ—Ė–Ĺ—É. –Ē–Ķ—Ź–ļ–į –į–ļ—ā–ł–≤–Ĺ—Ė—Ā—ā—Ć –į—Ä–≥—Ė–Ĺ–į–∑–ł —ā–į–ļ–ĺ–∂ —Ā–Ņ–ĺ—Ā—ā–Ķ—Ä—Ė–≥–į—Ē—ā—Ć—Ā—Ź —É –Ņ–ĺ–∑–į–ļ–Ľ—Ė—ā–ł–Ĺ–Ĺ–ł—Ö —Ä—Ė–ī–ł–Ĺ–į—Ö, –≤–ĺ–Ĺ–į –∑–Ĺ–į—á–Ĺ–ĺ –∑—Ä–ĺ—Ā—ā–į—Ē –≤ —É–ľ–ĺ–≤–į—Ö –∑–į–Ņ–į–Ľ–Ķ–Ĺ–Ĺ—Ź –į–Ī–ĺ —ā—Ä–į–≤–ľ[ 3] –ļ–ĺ–Ľ–į–≥–Ķ–Ĺ—É [ 5]
–ó–ĺ–≤—Ā—Ė–ľ –Ĺ–Ķ–ľ–į—Ē –į—Ä–≥—Ė–Ĺ–į–∑–ĺ–Ĺ–ĺ—ó –į–ļ—ā–ł–≤–Ĺ–ĺ—Ā—ā—Ė –≤ –ļ–Ľ—Ė—ā–ł–Ĺ–į—Ö –Ņ–Ľ–į—Ü–Ķ–Ĺ—ā–ł —Ā–≤–ł–Ĺ–Ķ–Ļ –Ļ –Ķ–Ĺ—ā–Ķ—Ä–ĺ—Ü–ł—ā–į—Ö –Ņ–ĺ—Ä–ĺ—Ā—Ź—ā, —Č–ĺ –≤–ł–≥–ĺ–ī–ĺ–≤—É—é—ā—Ć—Ā—Ź –ľ–ĺ–Ľ–ĺ–ļ–ĺ–ľ. –ó–į–≤–ī—Ź–ļ–ł —Ü—Ć–ĺ–ľ—É –ľ–į–ļ—Ā–ł–ľ–į–Ľ—Ć–Ĺ–į –ļ—Ė–Ľ—Ć–ļ—Ė—Ā—ā—Ć –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –ľ–ĺ–∂–Ķ –Ņ–ĺ—Ā—ā–į—á–į—ā–ł—Ā—Ć –≤—Ė–ī –ľ–į—ā–Ķ—Ä—Ė –ī–ĺ –ī–ł—ā–ł–Ĺ–ł. –ü—Ä–ĺ—ā–Ķ —ā–į–ļ–į –∑–į–ļ–ĺ–Ĺ–ĺ–ľ—Ė—Ä–Ĺ—Ė—Ā—ā—Ć —Ā–Ņ–ĺ—Ā—ā–Ķ—Ä—Ė–≥–į—Ē—ā—Ć—Ā—Ź –Ĺ–Ķ —É –≤—Ā—Ė—Ö –≤–ł–ī—Ė–≤, –Ĺ–į–Ņ—Ä–ł–ļ–Ľ–į–ī —ā—Ė –∂ —ā–ļ–į–Ĺ–ł–Ĺ–ł –ĺ–≤–Ķ—Ü—Ć –ľ–į—é—ā—Ć –ī–ĺ—Ā–ł—ā—Ć –≤–ł—Ā–ĺ–ļ—Ė —Ä—Ė–≤–Ĺ—Ė –į—Ä–≥—Ė–Ĺ–į–∑–ł[ 3] –Ľ–į–ļ—ā–į—Ü—Ė—ó , –ĺ—Ā–ļ—Ė–Ľ—Ć–ļ–ł –≤—Ė–Ĺ –Ĺ–Ķ–ĺ–Ī—Ö—Ė–ī–Ĺ–ł–Ļ –ī–Ľ—Ź —É—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź –Ņ—Ä–ĺ–Ľ—Ė–Ĺ—É —Ė–∑ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É[ 5]
–£ –Ľ—é–ī–ł–Ĺ–ł, —Č–ĺ –≤–į–∂–ł—ā—Ć 70 –ļ–≥ –Ņ—Ä–ł–Ī–Ľ–ł–∑–Ĺ–ĺ 2,3 –≥ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≤ –ī–Ķ–Ĺ—Ć –≤–ł—ā—Ä–į—á–į—Ē—ā—Ć—Ā—Ź –Ĺ–į —Ā–ł–Ĺ—ā–Ķ–∑ –ļ—Ä–Ķ–į—ā–ł–Ĺ—É (10 % –≤—Ā—Ć–ĺ–≥–ĺ –į—Ä–≥—Ė–Ĺ—Ė—É –≤ –ļ—Ä–ĺ–≤–ĺ–ĺ–Ī—Ė–≥—É). –¶—Ź —Ā–Ņ–ĺ–Ľ—É–ļ–į —É —Ą–ĺ—Ā—Ą–ĺ—Ä–ł–Ľ—Ć–≤–į–Ĺ–ĺ–ľ—É —Ā—ā–į–Ĺ—Ė (—Ą–ĺ—Ā—Ą–ĺ–ļ—Ä–Ķ–į—ā–ł–Ĺ ) —Ā–Ľ—É–≥—É—Ē —Ä–Ķ–∑–Ķ—Ä–≤—É–į—Ä–ĺ–ľ –≤—Ė–Ľ—Ć–Ĺ–ĺ—ó –Ķ–Ĺ–Ķ—Ä–≥—Ė—ó –ī–Ľ—Ź —ą–≤–ł–ī–ļ–ĺ—ó —Ä–Ķ–≥–Ķ–Ĺ–Ķ—Ä–į—Ü—Ė—ó –ź–Ę–§ —É —ā–ł—Ö —ā–ł–Ņ–į—Ö –ļ–Ľ—Ė—ā–ł–Ĺ, —Ź–ļ—Ė –Ļ–ĺ–≥–ĺ –ī—É–∂–Ķ —Ė–Ĺ—ā–Ķ–Ĺ—Ā–ł–≤–Ĺ–ĺ –≤–ł–ļ–ĺ—Ä–ł—Ā—ā–ĺ–≤—É—é—ā—Ć (–ľ'—Ź–∑–ł, –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–ł ). –ö—Ė–Ľ—Ć–ļ–ĺ—Ā—ā—Ė —Ą–ĺ—Ā—Ą–ĺ–ļ—Ä–Ķ–į—ā–ł–Ĺ—É –≤ —Ā–ļ–Ľ–Ķ—ā–Ķ–Ĺ–ł—Ö –ľ'—Ź–∑–į—Ö –Ľ—é–ī–ł–Ĺ–ł –ī–ĺ—Ā—ā–į—ā–Ĺ—Ć–ĺ –ī–Ľ—Ź –∑–į–Ī–Ķ–∑–Ņ–Ķ—á–Ķ–Ĺ–Ĺ—Ź —ó—Ö –Ķ–Ĺ–Ķ—Ä–≥—Ė—ó –≤–Ņ—Ä–ĺ–ī–ĺ–≤–∂ –ļ—Ė–Ľ—Ć–ļ–ĺ—Ö —Ö–≤–ł–Ľ–ł–Ĺ. –Ę–į–ļ–ĺ–∂ —Ē –≤—Ė–ī–ĺ–ľ–ĺ—Ā—ā—Ė –Ņ—Ä–ĺ —ā–Ķ, —Č–ĺ –ļ—Ä–Ķ–į—ā–ł–Ĺ –ľ–į—Ē –į–Ĺ—ā–ł–ĺ–ļ—Ā–ł–ī–į—ā–ł–≤–Ĺ—Ė —Ė –Ņ—Ä–ĺ—ā–ł–∑–į–Ņ–į–Ľ—Ć–Ĺ—Ė –≤–Ľ–į—Ā—ā–ł–≤–ĺ—Ā—ā—Ė, –į —ā–į–ļ–ĺ–∂ –Ņ–ĺ–ļ—Ä–į—Č—É—Ē –≥–Ľ—é–ļ–ĺ–∑–ĺ—ā–ĺ–Ľ–Ķ—Ä–į–Ĺ—ā–Ĺ—Ė—Ā—ā—Ć —É –Ľ—é–ī–Ķ–Ļ[ 3]
–§–ĺ—Ā—Ą–ĺ–ļ—Ä–Ķ–į—ā–ł–Ĺ —Ā–ł–Ĺ—ā–Ķ–∑—É—Ē—ā—Ć—Ā—Ź –≤ –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ—Ė –∑–į–≤–ī—Ź–ļ–ł –ľ–Ķ—ā–į–Ī–ĺ–Ľ—Ė—á–Ĺ—Ė–Ļ –ļ–ĺ–ĺ–Ņ–Ķ—Ä–į—Ü—Ė—ó –ľ—Ė–∂ –Ĺ–ł—Ä–ļ–į–ľ–ł, –Ņ—Ė–ī—ą–Ľ—É–Ĺ–ļ–ĺ–≤–ĺ—é –∑–į–Ľ–ĺ–∑–ĺ—é , –Ņ–Ķ—á—Ė–Ĺ–ļ–ĺ—é —ā–į —Ā–ļ–Ķ–Ľ–Ķ—ā–Ĺ–ł–ľ–ł –ľ'—Ź–∑–į–ľ–ł. –ü–Ķ—Ä—ą—É —Ä–Ķ–į–ļ—Ü—Ė—é –Ĺ–į —ą–Ľ—Ź—Ö—É –Ņ–Ķ—Ä–Ķ—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≤ –ļ—Ä–Ķ–į—ā–ł–Ĺ –ļ–į—ā–į–Ľ—Ė–∑—É—Ē –į—Ä–≥—Ė–Ĺ—Ė–Ĺ:–≥–Ľ—Ė—Ü–ł–Ĺ –į–ľ—Ė–ī–ł–Ĺ–ĺ—ā—Ä–į–Ĺ—Ā—Ą–Ķ—Ä–į–∑–į (–ö–§ 2.1.4.1) ‚ÄĒ –ľ—Ė—ā–ĺ—Ö–ĺ–Ĺ–ī—Ä—Ė–į–Ľ—Ć–Ĺ–ł–Ļ —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā, —Č–ĺ –Ķ–ļ—Ā–Ņ—Ä–Ķ—Ā—É—Ē—ā—Ć—Ā—Ź –≥–ĺ–Ľ–ĺ–≤–Ĺ–ł–ľ —á–ł–Ĺ–ĺ–ľ —É –Ĺ–ł—Ä–ļ–ĺ–≤–ł—Ö —ā—Ä—É–Ī–ĺ—á–ļ–į—Ö —ā–į –Ņ—Ė–ī—ą–Ľ—É–Ĺ–ļ–ĺ–≤—Ė–Ļ –∑–į–Ľ–ĺ–∑—Ė, —Ė –≤ –∑–Ĺ–į—á–Ĺ–ĺ –ľ–Ķ–Ĺ—ą—Ė–Ļ –ļ—Ė–Ľ—Ć–ļ–ĺ—Ā—ā—Ė –≤ –Ņ–Ķ—á—Ė–Ĺ—Ü—Ė —ā–į —Ė–Ĺ—ą–ł—Ö –ĺ—Ä–≥–į–Ĺ–į—Ö. –ü—Ä–ĺ–ī—É–ļ—ā–ĺ–ľ —Ü—Ė—Ē—ó —Ä–Ķ–į–ļ—Ü—Ė—ó, —Č–ĺ –Ņ–ĺ–Ľ—Ź–≥–į—Ē —É –Ņ–Ķ—Ä–Ķ–Ĺ–Ķ—Ā–Ķ–Ĺ–Ĺ—Ė –≥—É–į–Ĺ—Ė–ī–ł–Ĺ–ĺ–≤–ĺ—ó –≥—Ä—É–Ņ–ł —Ė–∑ –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –Ĺ–į –≥–Ľ—Ė—Ü–ł–Ĺ , —Ē –≥—É–į–Ĺ—Ė–ī–ł–Ĺ–ĺ–į—Ü–Ķ—ā–į—ā —ā–į –ĺ—Ä–Ĺ—Ė—ā–ł–Ĺ. –ď—É–į–Ĺ—Ė–ī–ł–Ĺ–ĺ–į—Ü–Ķ—ā–į—ā –Ņ—Ė–ī–Ľ—Ź–≥–į—Ē –ľ–Ķ—ā–ł–Ľ—é–≤–į–Ĺ–Ĺ—é —Ü–ł—ā–ĺ–Ņ–Ľ–į–∑–ľ–į—ā–ł—á–Ĺ–ł–ľ —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā–ĺ–ľ –≥—É–į–Ĺ—Ė–ī–ł–Ĺ–ĺ–į—Ü–Ķ—ā–į—ā-N-–ľ–Ķ—ā–ł–Ľ—ā—Ä–į–Ĺ—Ā—Ą–Ķ—Ä–į–∑–ĺ—é —Ė–∑ —É—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź–ľ –ļ—Ä–Ķ–į—ā–ł–Ĺ—É. –¶–Ķ–Ļ –Ķ—ā–į–Ņ –Ĺ–į–Ļ–Ī—Ė–Ľ—Ć—ą —Ė–Ĺ—ā–Ķ–Ĺ—Ā–ł–≤–Ĺ–ĺ –Ņ—Ä–ĺ—ā—Ė–ļ–į—Ē —É –Ņ–Ķ—á—Ė–Ĺ—Ü—Ė —ā–į –Ņ—Ė–ī—ą–Ľ—É–Ĺ–ļ–ĺ–≤—Ė–Ļ –∑–į–Ľ–ĺ–∑—Ė, —Ė –∑–Ĺ–į—á–Ĺ–ĺ –Ņ–ĺ–≤—Ė–Ľ—Ć–Ĺ—Ė—ą–Ķ —É –Ĺ–ł—Ä–ļ–į—Ö. –ö—Ä–Ķ–į—ā–ł–Ĺ –≤–ł–≤—Ė–Ľ—Ć–Ĺ—Ź—Ē—ā—Ć—Ā—Ź —É –ļ—Ä–ĺ–≤, –∑–≤—Ė–ī–ļ–ł –ľ–ĺ–∂–Ķ –Ņ–ĺ–≥–Ľ–ł–Ĺ–į—ā–ł—Ā—Ć –ľ'—Ź–∑–į–ľ–ł —Ė –Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–į–ľ–ł —ā–į —Ą–ĺ—Ā—Ą–ĺ—Ä–ł–Ľ—é–≤–į—ā–ł—Ā—Ć –ī–ĺ —Ą–ĺ—Ā—Ą–ĺ–ļ—Ä–Ķ–į—ā–ł–Ĺ—É[ 5]
–£ –ī–Ķ—Ź–ļ–ł—Ö –Ī–Ķ–∑—Ö—Ä–Ķ–Ī–Ķ—ā–Ĺ–ł—Ö , —ā–į–ļ–ł—Ö —Ź–ļ –ĺ–ľ–į—Ä , —Ą—É–Ĺ–ļ—Ü—Ė—é —Ą–ĺ—Ā—Ą–ĺ–ļ—Ä–Ķ–į—ā–ł–Ĺ—É –≤–ł–ļ–ĺ–Ĺ—É—Ē —Ą–ĺ—Ā—Ą–ĺ–į—Ä–≥—Ė–Ĺ—Ė–Ĺ .
–ź—Ä–≥—Ė–Ĺ—Ė–Ĺ —Ē –Ņ–ĺ–Ņ–Ķ—Ä–Ķ–ī–Ĺ–ł–ļ–ĺ–ľ –ī–Ľ—Ź —Ā–ł–Ĺ—ā–Ķ–∑—É –ľ–ĺ–Ĺ–ĺ–ĺ–ļ—Ā–ł–ī—É –į–∑–ĺ—ā—É (NO) ‚ÄĒ —Ā–ł–≥–Ĺ–į–Ľ—Ć–Ĺ–ĺ—ó –ľ–ĺ–Ľ–Ķ–ļ—É–Ľ–ł, —Č–ĺ –Ī–Ķ—Ä–Ķ —É—á–į—Ā—ā—Ć —É —Ä–Ķ–≥—É–Ľ—Ź—Ü—Ė—ó —Ä—Ź–ī—É —Ą—Ė–∑—Ė–ĺ–Ľ–ĺ–≥—Ė—á–Ĺ–ł—Ö –Ņ—Ä–ĺ—Ü–Ķ—Ā—Ė–≤, —ā–į–ļ–ł—Ö —Ź–ļ —Ä–ĺ–∑—ą–ł—Ä–Ķ–Ĺ–Ĺ—Ź –ļ—Ä–ĺ–≤–ĺ–Ĺ–ĺ—Ā–Ĺ–ł—Ö —Ā—É–ī–ł–Ĺ, –Ņ–Ķ—Ä–Ķ–ī–į—á–į –Ĺ–Ķ—Ä–≤–ĺ–≤–ł—Ö —Ė–ľ–Ņ—É–Ľ—Ć—Ā—Ė–≤ —ā–į —Ė–ľ—É–Ĺ–Ĺ–į –≤—Ė–ī–Ņ–ĺ–≤—Ė–ī—Ć . –£—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź NO –∑–į–Ī–Ķ–∑–Ņ–Ķ—á—É—Ē —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā NO-—Ā–ł–Ĺ—ā–į–∑–į, —Č–ĺ —É —Ā—Ā–į–≤—Ü—Ė–≤ –ľ–į—Ē —ā—Ä–ł —Ė–∑–ĺ—Ą–ĺ—Ä–ľ–ł (–Ĺ–Ķ–Ļ—Ä–ĺ–Ĺ–į–Ľ—Ć–Ĺ–į nNOS, –Ķ–Ĺ–ī–ĺ—ā–Ķ–Ľ—Ė–į–Ľ—Ć–Ĺ–į eNOS, —Ė–Ĺ–ī—É—Ü–ł–Ī–Ķ–Ľ—Ć–Ĺ–į iNOS), —Ź–ļ—Ė –≤—Ė–ī—Ä—Ė–∑–Ĺ—Ź—é—ā—Ć—Ā—Ź —Ą—É–Ĺ–ļ—Ü—Ė—Ź–ľ–ł, –Ņ–ĺ—ą–ł—Ä–Ķ–Ĺ–Ĺ—Ź–ľ —É —ā–ļ–į–Ĺ–ł–Ĺ–į—Ö —ā–į –ĺ—Ā–ĺ–Ī–Ľ–ł–≤–ĺ—Ā—ā—Ź–ľ–ł —Ä–Ķ–≥—É–Ľ—Ź—Ü—Ė—ó.
–†–Ķ–į–ļ—Ü—Ė—Ź –Ņ–Ķ—Ä–Ķ—ā–≤–ĺ—Ä–Ķ–Ĺ–Ĺ—Ź –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –ī–ĺ —Ü–ł—ā—Ä—É–Ľ—Ė–Ĺ—É —ā–į NO –≤—Ė–ī–Ī—É–≤–į—Ē—ā—Ć—Ā—Ź —É –ī–≤–į –Ķ—ā–į–Ņ–ł —Ė–∑ –Ņ—Ä–ĺ–ľ—Ė–∂–Ĺ–ł–ľ –Ņ—Ä–ĺ–ī—É–ļ—ā–ĺ–ľ NŌČ -–≥—Ė–ī—Ä–ĺ–ļ—Ā–ł–Ľ-l -–į—Ä–≥—Ė–Ĺ—Ė–Ĺ (NOHA) —ā–į –Ņ–ĺ—ā—Ä–Ķ–Ī—É—Ē –ļ–ł—Ā–Ĺ—é, –Ě–ź–Ē–§–Ě, 5,6,7,8-—ā–Ķ—ā—Ä–į–≥—Ė–ī—Ä–ĺ–Ī—Ė–ĺ–Ņ—ā–Ķ—Ä–ł–Ĺ—É, –§–ú–Ě —ā–į –§–ź–Ē.
–§–Ķ—Ä–ľ–Ķ–Ĺ—ā –į—Ä–≥—Ė–Ĺ—Ė–Ĺ–ī–Ķ–ļ–į—Ä–Ī–ĺ–ļ—Ā–ł–Ľ–į–∑–į , —Ź–ļ–ł–Ļ –ļ–į—ā–į–Ľ—Ė–∑—É—Ē –ī–Ķ–ļ–į—Ä–Ī–ĺ–ļ—Ā–ł–Ľ—é–≤–į–Ĺ–Ĺ—Ź –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –ī–ĺ –į–≥–ľ–į—ā–ł–Ĺ—É, –Ī—É–≤ —Ā–Ņ–Ķ—Ä—ą—É –≤–ł—Ź–≤–Ľ–Ķ–Ĺ–ł–Ļ —É —Ä–ĺ—Ā–Ľ–ł–Ĺ–Ĺ–ł—Ö —ā–į –Ī–į–ļ—ā–Ķ—Ä—Ė–Ļ–Ĺ–ł—Ö –ļ–Ľ—Ė—ā–ł–Ĺ–į—Ö, –Ņ—Ä–ĺ—ā–Ķ –Ņ—Ė–∑–Ĺ—Ė—ą–Ķ –Ļ–ĺ–≥–ĺ –∑–Ĺ–į–Ļ—ą–Ľ–ł —Ė –≤ –ľ–ĺ–∑–ļ—É, –Ņ–Ķ—á—Ė–Ĺ—Ü—Ė, –Ĺ–ł—Ä–ļ–į—Ö, –Ĺ–į–ī–Ĺ–ł—Ä–ļ–ĺ–≤–ł—Ö –∑–į–Ľ–ĺ–∑–į—Ö, –ľ–į–ļ—Ä–ĺ—Ą–į–≥–į—Ö —ā–į —ā–ĺ–Ĺ–ļ—Ė–Ļ –ļ–ł—ą—Ü—Ė —Ā—Ā–į–≤—Ü—Ė–≤ . –§—Ė–∑—Ė–ĺ–Ľ–ĺ–≥—Ė—á–Ĺ–į —Ä–ĺ–Ľ—Ć –į–≥–ľ–į—ā–ł–Ĺ—É —ā–ĺ—á–Ĺ–ĺ –Ĺ–Ķ –≤—Ā—ā–į–Ĺ–ĺ–≤–Ľ–Ķ–Ĺ–į, –Ņ—Ä–ĺ—ā–Ķ —ā—Ä–ł–≤–į—é—ā—Ć –Ļ–ĺ–≥–ĺ –ī–ĺ—Ā–Ľ—Ė–ī–∂–Ķ–Ĺ–Ĺ—Ź —Ź–ļ –ľ–ĺ–∂–Ľ–ł–≤–ĺ–≥–ĺ –Ĺ–Ķ–Ļ—Ä–ĺ–ľ–Ķ–ī—Ė–į—ā–ĺ—Ä–į —ā–į —Ä–Ķ–≥—É–Ľ—Ź—ā–ĺ—Ä–į –Ī–į–≥–į—ā—Ć–ĺ—Ö —Ą—É–Ĺ–ļ—Ü—Ė–Ļ –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ—É[ 5] [ 22]
–£ –ī–Ķ—Ź–ļ–ł—Ö –Ľ—Ė–Ĺ—Ė—Ź—Ö –∑–Ľ–ĺ—Ź–ļ—Ė—Ā–Ĺ–ł—Ö –ļ–Ľ—Ė—ā–ł–Ĺ, –∑–ĺ–ļ—Ä–Ķ–ľ–į –ľ–Ķ–Ľ–į–Ĺ–ĺ–ľ–ł , –≥–Ķ–Ņ–į—ā–ĺ—Ü–Ķ–Ľ—é–Ľ—Ź—Ä–Ĺ–ĺ—ó –ļ–į—Ä—Ü–ł–Ĺ–ĺ–ľ–ł , –Ĺ–ł—Ä–ļ–ĺ–≤–ĺ–ļ–Ľ—Ė—ā–ł–Ĺ–Ĺ–ĺ–≥–ĺ —Ä–į–ļ—É —ā–į –ī–Ķ—Ź–ļ–ł—Ö –ľ–Ķ–∑–ĺ—ā–Ķ–Ľ—Ė–ĺ–ľ , —Ā–Ņ–ĺ—Ā—ā–Ķ—Ä—Ė–≥–į—Ē—ā—Ć—Ā—Ź –Ī—Ä–į–ļ –Ķ–ļ—Ā–Ņ—Ä–Ķ—Ā—Ė—ó –į—Ä–≥—Ė–Ĺ—Ė–Ĺ–ĺ—Ā—É–ļ—Ü–ł–Ĺ–į—ā—Ā–ł–Ĺ—ā–į–∑–ł, –≤–Ĺ–į—Ā–Ľ—Ė–ī–ĺ–ļ —á–ĺ–≥–ĺ –≤–ĺ–Ĺ–ł —Ā—ā–į—é—ā—Ć –į—É–ļ—Ā–ĺ—ā—Ä–ĺ—Ą–Ĺ–ł–ľ–ł –∑–į –į—Ä–≥—Ė–Ĺ—Ė–Ĺ–ĺ–ľ. –ß–Ķ—Ä–Ķ–∑ —Ü–Ķ —Ą–Ķ—Ä–ľ–Ķ–Ĺ—ā–ł, —Č–ĺ —Ä–ĺ–∑—Č–Ķ–Ņ–Ľ—é—é—ā—Ć –į—Ä–≥—Ė–Ĺ—Ė–Ĺ, —ā–į–ļ—Ė —Ź–ļ –į—Ä–≥—Ė–Ĺ–į–∑–į —ā–į –Ī–į–ļ—ā–Ķ—Ä—Ė–Ļ–Ĺ–į –į—Ä–≥—Ė–Ĺ—Ė–Ĺ–ī–Ķ—Ė–ľ—Ė–Ĺ–į–∑–į , –ľ–ĺ–∂—É—ā—Ć –≤–ł–ļ–ĺ—Ä–ł—Ā—ā–ĺ–≤—É–≤–į—ā–ł—Ā—Ć —Ź–ļ –Ņ—Ä–ĺ—ā–ł–Ņ—É—Ö–Ľ–ł–Ĺ–Ĺ—Ė –Ņ—Ä–Ķ–Ņ–į—Ä–į—ā–ł –Ņ—Ä–ł —Ü–ł—Ö —ā–ł–Ņ–į—Ö —Ä–į–ļ—É[ 23]
–í–ľ—Ė—Ā—ā –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≤—Ė–ī–Ĺ–ĺ—Ā–Ĺ–ĺ –≤–ł—Ā–ĺ–ļ–ł–Ļ —É –ľ–ĺ—Ä—Ā—Ć–ļ—Ė–Ļ —ó–∂—Ė, —Ā–ĺ–ļ—É –ļ–į–≤—É–Ĺ–į, –≥–ĺ—Ä—Ė—Ö–į—Ö, –Ĺ–į—Ā—Ė–Ĺ–Ĺ—Ė, –≤–ĺ–ī–ĺ—Ä–ĺ—Ā—ā—Ź—Ö, –ľ'—Ź—Ā—Ė, –ļ–ĺ–Ĺ—Ü–Ķ–Ĺ—ā—Ä–į—ā—Ė —Ä–ł—Ā–ĺ–≤–ĺ–≥–ĺ –Ī—Ė–Ľ–ļ–į —ā–į —É —Ā–ĺ—Ē–≤–ĺ–ľ—É –Ī—Ė–Ľ–ļ—É, –į–Ľ–Ķ –Ĺ–ł–∑—Ć–ļ–ł–Ļ —É –ľ–ĺ–Ľ–ĺ—Ü—Ė –Ī—Ė–Ľ—Ć—ą–ĺ—Ā—ā—Ė —Ā—Ā–į–≤—Ü—Ė–≤ (–≤–ļ–Ľ—é—á–Ĺ–ĺ —Ė–∑ –Ľ—é–ī–ł–Ĺ–ĺ—é, –ļ–ĺ—Ä–ĺ–≤–į–ľ–ł —Ė —Ā–≤–ł–Ĺ—Ć–ľ–ł). –ü—Ä–ĺ—ā–Ķ –≤–Ķ–Ľ–ł–ļ–į —á–į—Ā—ā–ł–Ĺ–į –≤—Ė–ī –∑–į–≥–į–Ľ—Ć–Ĺ–ĺ—ó –ļ—Ė–Ľ—Ć–ļ–ĺ—Ā—ā—Ė —Ü—Ė—Ē—ó –į–ľ—Ė–Ĺ–ĺ–ļ–ł—Ā–Ľ–ĺ—ā–ł, —Č–ĺ –Ņ–ĺ—Ā—ā—É–Ņ–į—Ē –≤ –ĺ—Ä–≥–į–Ĺ—Ė–∑–ľ –ī–ĺ—Ä–ĺ—Ā–Ľ–ĺ—ó –Ľ—é–ī–ł–Ĺ–ł —Ė–∑ —ó–∂–Ķ—é, –Ĺ–Ķ –Ņ–ĺ—ā—Ä–į–Ņ–Ľ—Ź—Ē –≤ –ļ—Ä–ĺ–≤, –ĺ—Ā–ļ—Ė–Ľ—Ć–ļ–ł –Ī–Ľ–ł–∑—Ć–ļ–ĺ 40 % –į—Ä–≥—Ė–Ĺ—Ė–Ĺ—É –≤—Ė–ī—Ä–į–∑—É –∂ —Ä–ĺ–∑—Č–Ķ–Ņ–Ľ—é—Ē—ā—Ć—Ā—Ź —É —ā–ĺ–Ĺ–ļ—Ė–Ļ –ļ–ł—ą—Ü—Ė[ 3]
‚ÜĎ –į –Ī –≤ –≥ –ī –Ķ –∂ Wu G, Bazer FW, Davis TA, Kim SW, Li P, Marc Rhoads J, Carey Satterfield M, Smith SB, Spencer TE, Yin Y (2008). Arginine metabolism and nutrition in growth, health and disease. Amino Acids . 37 : 153‚ÄĒ68. doi :10.1007/s00726-008-0210-y . PMID 19030957 . ‚ÜĎ –į –Ī Morris SM Jr (2006). Arginine: beyond protein . Am J Clin Nutr . 83 : 508S‚ÄĒ512S. PMID 16470022 . –ź—Ä—Ö—Ė–≤ –ĺ—Ä–ł–≥—Ė–Ĺ–į–Ľ—É –∑–į 19 –≥—Ä—É–ī–Ĺ—Ź 2011. –ü—Ä–ĺ—Ü–ł—ā–ĺ–≤–į–Ĺ–ĺ 14 —Ā–Ķ—Ä–Ņ–Ĺ—Ź 2012 . ‚ÜĎ –į –Ī –≤ –≥ –ī –Ķ –∂ –ł –ļ –Ľ –ľ –Ĺ –Ņ —Ä —Ā —ā —É —Ą Wu G, Morris SM Jr (1998). Arginine metabolism: nitric oxide and beyond . Biochem J . 15 : 1‚ÄĒ17. PMID 9806879 . –ź—Ä—Ö—Ė–≤ –ĺ—Ä–ł–≥—Ė–Ĺ–į–Ľ—É –∑–į 15 —Ā–Ķ—Ä–Ņ–Ĺ—Ź 2002. –ü—Ä–ĺ—Ü–ł—ā–ĺ–≤–į–Ĺ–ĺ 14 —Ā–Ķ—Ä–Ņ–Ĺ—Ź 2012 . ‚ÜĎ Jorpes E, Thor√©n S (1932). The use of the Sakaguchi reaction for the quantitative determination of arginine (PDF) . Biochem J . 26 : 1504‚ÄĒ6. PMID 16744969 . ‚ÜĎ Vossenaar ER, Zendman AJ, van Venrooij WJ, Pruijn GJ (2003). PAD, a growing family of citrullinating enzymes: genes, features and involvement in disease. Bioessays . 25 : 1106‚ÄĒ18. doi :10.1002/bies.10357 . PMID 14579251 . ‚ÜĎ Galgano F, Caruso M, Condelli N, Favati F (2012). Focused review: agmatine in fermented foods. Front Microbiol . 3 . doi :10.3389/fmicb.2012.00199 . PMID 22701114 . {{cite journal }}: –ě–Ī—Ā–Ľ—É–≥–ĺ–≤—É–≤–į–Ĺ–Ĺ—Ź CS1: –°—ā–ĺ—Ä—Ė–Ĺ–ļ–ł —Ė–∑ –Ĺ–Ķ–Ņ–ĺ–∑–Ĺ–į—á–Ķ–Ĺ–ł–ľ DOI –∑ –Ī–Ķ–∑–ļ–ĺ—ą—ā–ĺ–≤–Ĺ–ł–ľ –ī–ĺ—Ā—ā—É–Ņ–ĺ–ľ (–Ņ–ĺ—Ā–ł–Ľ–į–Ĺ–Ĺ—Ź )‚ÜĎ Feun L, You M, Wu CJ, Kuo MT, Wangpaichitr M, Spector S, Savaraj N (2008). Arginine deprivation as a targeted therapy for cancer. Curr Pharm Des . 14 : 1049‚ÄĒ57. PMID 18473854 .





















")

