![]() ヒト(男性)の染色体構造におけるX染色体
ヒト(男性)の染色体構造におけるX染色体
X染色体(エックスせんしょくたい)とは有性生殖をする真核生物にみられる性染色体の一種である。雌が性染色体として相同染色体の対を持つとき、それをX染色体と呼ぶ。このとき、雄はX染色体と共にY染色体を組として持つか(XY型)、あるいは対にならないX染色体のみを持つ(XO型)。このような性決定様式は雄がヘテロ型であるため「雄ヘテロ型」と呼ぶ。
概論
表. X染色体数と常染色体セット数の
比率(X/A)での性決定の例[1][2]
| 性 |
X/A |
常染色体
セット数(A) |
X染色体
数(X)
|
| 雄 |
0.33 |
3 |
1
|
| 0.50 |
2 |
1
|
| 0.50 |
4 |
2
|
| 間性 |
0.67 |
3 |
2
|
| 0.75 |
4 |
3
|
| 0.86 |
7 |
6
|
| 雌 |
1.00 |
2 |
2
|
| 1.00 |
3 |
3
|
| 1.00 |
4 |
4
|
|
| 小野知夫「高等植物の性決定と分化」(『最近の生物学』第4巻, 1951年)37ページの表「スイバの染色体組合せと性型」、東京農工大学農学部蚕学研究室『性決定』5ページの表「キイロショウジョウバエの染色体構成と性」より作成。「キイロショウジョウバエの染色体構成と性」の元データはFrost(1960), Goldscmidt(1955)による。
|
| Y染色体数は性決定に関係が無いので省略した。 X/A ≧ 1.0 では雌、1.0> X/A >0.5では間性、0.5 ≧ X/Aでは雄になる。
|
性決定
ヒトを含む哺乳類の大部分はXY型性決定方式である。他にショウジョウバエなど昆虫の一部や植物のヒロハノマンテマ・スイバなどがXY型であることが知られている。XO型の性決定様式の代表例は、線形動物Caenorhabditis elegansおよび一部の昆虫(バッタ・カメムシ)などがある。
雄ヘテロ型の性決定では、哺乳類およびヒロハノマンテマのようにY染色体の存在が雄への性分化を決定する例と、ショウジョウバエおよびスイバのようにX染色体の数と常染色体のセット数の比率(X/A)で性決定する例とがある。この違いは、前者ではY染色体の雄性化因子(哺乳類ではSRY遺伝子)で性決定するのに対して、後者はY染色体の有無・本数に関わらずX染色体の数に依存して性別が定まる(右表)。ショウジョウバエでは、ニューメレーター因子(numerator elements, 計数因子)がX染色体の数を認識していると考えられている[3]。
由来と構造
X染色体とY染色体は、1対の常染色体が分化して生じ、特定の生物グループごとに独立に成立したと考えられている。例えば、哺乳類の中でもカモノハシなどの単孔類は他の哺乳類よりも鳥類の性染色体と配列類似性が高く、単孔類とそれ以外の哺乳類(獣亜綱)の性染色体は別の起原を持つ[4]。また、メダカ・ヒロハノマンテマのX染色体およびY染色体も、それぞれの属が成立した後に、属内の種分化によって成立したと考えられている[5][6]。
X染色体は、雌においてはホモ型となり相同染色体の間での組換え[注釈 1]を起こすことができるため、突然変異などの影響を比較的に受けにくく、遺伝情報を維持しやすい。X染色体の遺伝情報量が維持されてきたという仮説は、1967年に大野乾によって提唱されている(大野の法則)[7]。
X染色体とY染色体は、進化の過程で相同性を失うように変化してきたが、相同性が残っている領域もある。常染色体のようにX染色体とY染色体の間で相同乗り換えが起きるため、その領域は擬似常染色体領域(PAR, pseudoautosomal region)と呼ばれている。
研究史
X染色体はヘルマン・ヘンキングによって1891年に発見された。ヘンキングは、この染色体の特殊性について充分には気づかず「X染色体」と名付けて発表した[8]。その後、1900年代に染色体研究が進展し、X染色体が性決定に関与する染色体であることが判明した[9]。X染色体という名称はヘンキングの命名によるものであり「正体不明」の意味と伝えられる。X染色体・Y染色体がそれぞれアルファベットのX・Yのような形をしているからそう呼ばれるようになったというのは俗説である[10]。
1949年にマレー・バーが見つけた哺乳類の雌特異的な細胞内構造物は、一般にバー小体と呼ばれた。後に、これがX染色体がヘテロクロマチン構造をとるように変化したものであることが示され、哺乳類での遺伝子量補償に関わっていることが判明した[11]。
ヒトのX染色体
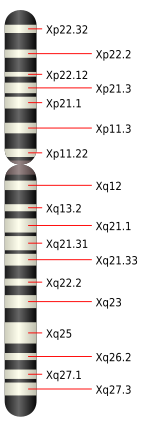 ヒトのX染色体模式図
ヒトのX染色体模式図
上部が短腕、くびれがセントロメア、下部が長腕である。上端の黒く示した部分(Xp22.33)の一部が擬似常染色体領域1(PAR1)である。最下端の擬似常染色体領域2(PAR2)は縮尺上表示できないほど小さいので省略してある。血友病の遺伝子はXq26.3(Xq26.2とXq27.1の間)、色覚関連(赤オプシン・緑オプシン)遺伝子は長腕末端部の黒く示した部分(Xq28)に存在する。
以下にヒトのX染色体について解説するが、他の生物では該当しない情報も含まれていることには注意が必要である。
構成
ヒトのX染色体はおよそ1億6,300万塩基対であり、7番染色体とほぼ同じ大きさを持つ。含まれる遺伝子の数は、資料によって異なるが、1,098個とする報告がある[12]。
X染色体には生命維持に欠かせない遺伝子が含まれており[13]、神経細胞のネットワーク構築・情報伝達、免疫機能関連遺伝子、(異常が起きると血友病となる)血液凝固に関連する遺伝子が例として挙げられる[14]。それらの多くはX染色体独自の領域に存在し、伴性遺伝をする。
短腕末端部分と長腕末端部分にはそれぞれY染色体と相同な領域である擬似常染色体領域が存在する。
伴性遺伝
性染色体に存在する遺伝子の発現により、雌雄・男女で差が生じる遺伝のことを伴性遺伝という。一般的には血友病・色覚異常などの例が良く知られている。一般的に伴性遺伝疾患について次のように説明される[15]。
X染色体の特定遺伝子に異常がある確率をp(0<p<1)とする。このとき、男性はX染色体が1本しかないので、確率pでそのまま
表現型に現れる。女性はX染色体が2本あるので、両方のX染色体に同じ異常が揃う確率はp
2となり、より低い確率でしか表現型に現れない。仮にpに1,000分の1を代入すると、男性は1,000人に1人、女性は100万人に1人となり、大きな差ができる。
これは概論としては正しいが、厳密には正確性を欠く説明である。哺乳類の雌の体細胞では、2本のX染色体のうち片方が不活性化されている。ヒトを含む真獣下綱動物では、この不活性化がランダムに起き、雌の体細胞は2種類のX染色体のどちらが働いているかについて、モザイク状態になっている。したがって、疾患遺伝子をヘテロ接合型に持つ女性(保因者(英語版))では、細胞によっては遺伝子疾患が発症していることがある[16]。部分的に発症しているにもかかわらず、保因者女性で伴性遺伝疾患が強く現れないのは、モザイクを作っている正常な細胞による補完によるものであるとされている[17][18]。
X染色体に免疫系で働く遺伝子が含まれていることで、「女性が男性よりも強い免疫力を持っている」という説もある。しかしながら、それらの遺伝子の多くはX染色体の不活性化により遺伝子量補償(男女・雌雄の遺伝子発現量の均等化)がなされていると考えられることから、この説には確証があるわけではない。
性染色体数異常
性染色体数の異常によって、性染色体がX染色体1本のみである女性(ターナー症候群)、X染色体が過剰な男性(クラインフェルター症候群)などが生じる。詳しくは染色体異常#性染色体の異常を参照のこと。
脚注
注釈
- ^ (遺伝的)組換え - 遺伝子の組合せが入れ替わること。同じ現象を染色体の間の可視現象として捉えると染色体の乗換え(染色体交差)となる。
出典
- ^ 小野知夫「高等植物の性決定と分化」(『最近の生物学』第4巻)
- ^ 東京農工大学農学部蚕学研究室『性決定』
- ^ Cline TW (1988). “Evidence that sisterless-a and sisterless-b are two of several discrete "numerator elements" of the X/A sex determination signal in Drosophila that switch Sxl between two alternative stable expression states” (pdf). Genetics 119: 829-862. PMID 3137120. http://www.genetics.org/cgi/reprint/119/4/829.pdf 2009年5月2日閲覧。.
- ^ Veyrunes F, Waters PD, Miethke P, Rens W, McMillan D, Alsop AE, Grützner F, Deakin JE, Whittington CM, Schatzkamer K, Kremitzki CL, Graves T, Ferguson-Smith MA, Warren W, Marshall Graves JA (2008). “Bird-like sex chromosomes of platypus imply recent origin of mammal sex chromosomes”. Genome Res 18: 965-973. doi:10.1101/gr.7101908. http://genome.cshlp.org/content/18/6/965.full 2009年5月2日閲覧。.
- ^ 大竹博之、酒泉満「メダカ性決定遺伝子の起源と進化」『生物の科学 遺伝』2009年1月号
- ^ 松永幸大「高等植物の性決定機構」『蛋白質核酸酵素』2000年7月号
- ^ Ohno S (1967). Sex Chromosomes and sex-linked genes. Berlin:Springer-Verlag.
- ^ デイヴィッド・ベインブリッジ『X染色体:男と女を決めるもの』12-15ページ。原著論文は Henking, H(1891). L. Zeit. Wiss. Zool. 51.
- ^ 『X染色体:男と女を決めるもの』21-28ページ。原著論文は McLung CE(1902) Biological Bulletin 3: 43; McLung CE(1901) Anatomischer Anzeinger 20: 220; Steven NM(1905). Journal of Experimental Zoology 2: 371; Wilson EB(1905). Science 22: 500.
- ^ 「性を決めるカラクリ『X・Y染色体』」『Newton』、58ページ
- ^ 『X染色体:男と女を決めるもの』182-185ページ。
- ^ 「性を決めるカラクリ『X・Y染色体』」『Newton』、34-35ページ。
- ^ 「性を決めるカラクリ『X・Y染色体』」『Newton』、60-61ページ。
- ^ 「性を決めるカラクリ『X・Y染色体』」『Newton』、44-49ページ。
- ^ 「性を決めるカラクリ『X・Y染色体』」『Newton』、48-49ページ。
- ^ Puck J (1998). “X inactivation in females with X-linked disease”. NEJM. 338: 325-328. PMID 9445416.
- ^ 「性を決めるカラクリ『X・Y染色体』」『Newton』、50-53ページ。
- ^ デイヴィッド・ベインブリッジ『X染色体:男と女を決めるもの』201-230ページ
参考文献
- デイヴィッド・ベインブリッジ『X染色体:男と女を決めるもの』長野敬、小野木明恵(翻訳)、青土社 、2004年、ISBN 978-4791761524。
- 松永幸大(2000年)「高等植物の性決定機構」『蛋白質核酸酵素』2000年7月号(Vol.45 No.10)、共立出版、1704-1712ページ。2009-04-02閲覧。
- 諸橋憲一郎 他「性を決めるカラクリ,『X・Y染色体』」、『Newton』2006年2月号、2006年、ニュートンプレス。
- 大竹博之、酒泉満(2009年)「メダカ性決定遺伝子の起源と進化」『生物の科学 遺伝』2009年1月号、44-48ページ、TNS。
- 小野知夫「高等植物の性決定と分化」(駒井卓、木原均編『最近の生物学』第4巻)培風館、30-47ページ、1951年。
- 東京農工大学農学部蚕学研究室『昆虫の性染色体』『性決定』 2009-03-30 閲覧。
関連項目

