'白い体色を持つオオシモフリエダシャク Biston betularia f. typica' 黒い体色を持つBiston betularia f. carbonaria 工業暗化 (こうぎょうあんか、英 : industrial melanism )は、「19世紀後半から、ヨーロッパの工業都市が発展するにつれて、その付近に生息するガ(蛾 )に暗色の変異が増加した[ 1]
「工業暗化」の例としては、しばしばオオシモフリエダシャク (英語版 ) Biston betularia )が言及される[ 1] f.typica )、明るい色をした木や苔の上では効果的な保護色 となっていたが、19世紀半ばごろから黒い色の個体(暗化型;イギリスのものはf.carbonaria )が報告され始め、一部の個体群ではほとんどを暗化型が占めるに至った。最初に注目されたのはイギリスであったが、その後はヨーロッパの各地で見られるようになった[ 1] 産業革命 の初期にあたり、イギリス のロンドンからマンチェスターにかけての地域では、石炭を燃やすことによって生じる大気汚染 のため、コケのほとんどが死に絶え、木の樹皮はすす で黒く汚れていた[ 2] [ 2] 自然選択説 ベースの説明がされるようになる[ 2]
そして環境汚染に応じて個体群が暗化(黒化)することを指す「工業暗化 」の語が作り出された。この現象は単純でわかりやすく、オオシモフリエダシャクは自然選択 による進化を説明したり例示するときに使われる代表的な例となった[ 3] [ 1]
オオシモフリエダシャクの工業暗化の主要因が、鳥による蛾の捕食に起因する自然選択であることは、様々な実験や観察データの積み重ねから実証されている[ 4] [ 5] [ 6] [ 7] [ 8] [ 5] [ 9] [ 2]
なお、蛾以外の生物の体色が暗化した現象についても、この工業暗化の説明やそれに類似した説明が持ち出されることがあるが、そうした説明は必ずしも正しいとは限らない[ 1]
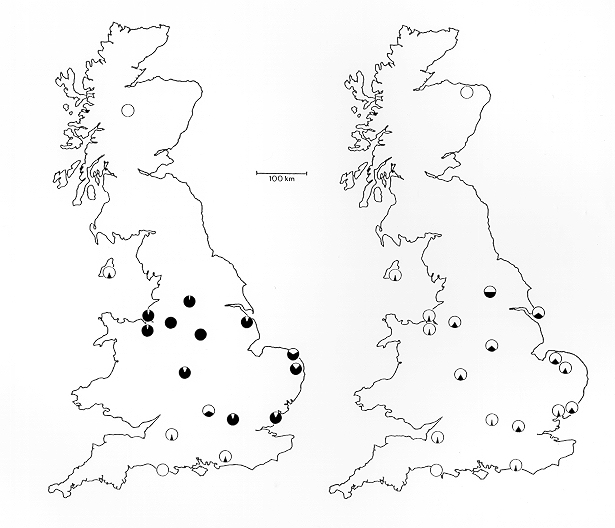
汚染されていない木では白い個体の方がうまくカモフラージュされており、捕食から逃れやすい。 暗化型の増加はヨーロッパと北アメリカのオオシモフリエダシャク個体群でおきた。暗化型個体の割合の増加に関する情報は乏しいが、その後の暗化型個体の割合の低下については、より詳しく調べられている。
産業革命 の前は、オオシモフリエダシャクのほとんどは、明るい地にわずかな黒いまだら模様を持つ淡色型ばかりだった。黒いオオシモフリエダシャク個体の最初の標本は、1811年以前の由来不明のものである[ 10] f. carboniana )と見なせる最初の個体は、1848年にマンチェスターで捕獲されたものである[ 2] [ 11] [ 11] [ 12] [ 12]
現代ではヨーロッパと北アメリカのより厳しい大気環境基準によって、汚染は減少し、それにともなって、1962年から現在に至るまで暗化型の頻度は着実に減少している。暗化型個体の減少は、科学研究の厳格化もあり、増加したときよりも正確に調べられた。特にバーナード・ケトルウェル( en:Bernard Kettlewell )は1956年に全国的な調査を行った。1996年には同様の調査をブルース・グラントが[ 13] [ 14]
同様の結果はアメリカでも観察された。日本では暗化型の蛾は見つかっていない。これは日本のエダシャク蛾が工業化された地域に生息していないからだと考えられている[ 14]
J.W.タット (英語版 ) [ 15] 葉状地衣類 )に被われていない黒い幹の上では優れたカモフラージュとなる。一方、明るい個体はコケに被われた白い幹の上で優れたカモフラージュとなる。結果として、鳥はうまくカモフラージュしていないほう個体をより頻繁に見つけて食べることになる[ 16] [ 16]
オオシモフリエダシャクの工業暗化の要因について、鳥の捕食による自然選択以外の要因を主張する代替仮説は主に1920年代から1930年代にかけて提案された。
ジョン・ウィリアム・ヘスロップ=ハリソン (1920)はタットの捕食仮説を否定した。そして代わりに汚染物質が生物の体細胞と生殖細胞の変化の原因となり得るという仮説を提唱した。この仮説はおそらく1890年代に提唱されていたラマルキズム の一種にルーツを持っている。歴史的な文脈に注意することは重要である。
ヘイズブローク(1925)は最初にこの仮説の検証を試みた。彼は大気汚染がチョウ類の生理に影響を与え、黒い色素の過剰生成を引き起こすと主張した。彼は蛾のまゆを様々な種類の汚染ガス(たとえば硫化水素 、アンモニア 、ピリジン )に曝した。その研究で8つの種が使われた。そのうち暗化しなかった4つの種はチョウだった。フォード(1964)はヘイズブロークの例で示された変異型は、暗化を示しておらず、またヘイズブロークは遺伝学を理解できなかったと主張した。
ヘスロップ=ハリソン(Harrison&Garrett 1926, Harrison 1928)は工業地帯の暗化型個体の増加は捕食による選択のためでなく、「突然変異圧」のためであると主張した。彼は、大気中の汚染粒子に含まれていた鉛やマンガンの塩がメラニン生成の遺伝子に変異を引き起こし、それ以外が引き起こしたのではないと主張した。ヘスロップ=ハリソンはSelenia bilunaria とTephrosia bistortata を用いた。幼虫にこれらの塩を含んだ葉を食べさせると暗化が起きた。だが同様の実験を行ったヒューズ・マッケンニー(1932)やトマセンとレムシェ(1933)はこの現象を再現することができなかった。統計学者で遺伝学者のロナルド・フィッシャー はヘスロップ=ハリソンの操作が不適切であったことを示した[ 17]
交配実験から、オオシモフリエダシャクの暗化型個体か淡色型個体かは遺伝的なものであり、暗化型個体か淡色型個体かを決めるのは一つの遺伝子座上にある対立遺伝子であると推定された[ 18] f.carboniana )とアメリカの暗化型個体(f.swettaria )について、遺伝的分析と交雑実験は、どちらも暗化型の表現型は常染色体 の顕性な形質として受け継がれ、これらの表現型は同一の遺伝子座 にあるアイソアレル(isoallele;別々の対立遺伝子)によって生み出されていることを示している[ 19] 平行進化 のモデルと言える。
近年の研究で、イギリスにおける黒色個体(f.carboniana )を生み出す対立遺伝子は、17番染色体のcortex遺伝子の最初のイントロンに生じた転移因子 の挿入によって突然変異として生じたことが示唆されている[ 20]
食虫性の鳥、シジュウカラ
遺伝学においては、進化の定義の一つは「遺伝子プール における対立遺伝子 の頻度の変化」であり[ 21] 個体群 の特徴に、変化が起きることである。集団遺伝学からは、遺伝子頻度の変化が起きなければ進化が起きないことがわかる。それぞれの遺伝子頻度をpとqで表し、たとえばp=0.6でq=0.4とする。そして個体群サイズが200から100まで減った場合でも、両者が同じだけ捕食されたとすると遺伝子頻度はp=0.6、q=0.4のままである。遺伝子頻度の変化を引き起こす要因としては、自然選択 、突然変異 、他地域からの移住、遺伝的浮動 が挙げられる[ 21]
1924年にJ・B・S・ホールデン は単純な一般選択モデル(general selection model)を用いて、観察されたようなオオシモフリエダシャクの進化に必要な選択上の有利さを計算した。彼は1848年に暗色個体の割合が2%で、1895年には95%になったと仮定し、その結果、暗色個体は明るい個体の1.5倍の有利さ(選択係数0.3以上)を持っていたことがわかった。モデルの問題を考慮しても、変化の速度があまりに速かったため、遺伝的浮動の可能性は合理的に除外された[ 22]
なお、オオシモフリエダシャクでは黒色個体を生み出す対立遺伝子が顕性であり、黒色表現型の個体の遺伝子型はホモ接合およびヘテロ接合、白色表現型の個体の遺伝子型はホモ接合のみとなるが、このことは個体群の暗化がなぜそれほど急激に起きたかを説明する助けとなる。
オオシモフリエダシャクの実験でもっとも有名なのは、E.B.フォード の指導下で行われたバーナード・ケトルウェル (英語版 ) シジュウカラ についばまれた。 1953年にバーミンガム のCadbury Nature Reserveで行われた実験では、エダシャクはマーキングされて放たれ、その後再捕獲された。彼はこの汚染された森で淡色型個体が優先的に捕食されていることに気づいた。このようにケトルウェルは汚染された生息地では、暗化型であることが生き残りに重要であることを示した。1955年にはドーセット州 の汚染されていない森で、それから再びバーミンガムの汚染林で、同様の実験を繰り返した。またニコ・ティンバーゲン を伴って実験を行い、共同でフィルムを撮った。他の研究者の調査でも同様の結果が明らかとなり、1996年にはヨーロッパと北アメリカで暗化個体の頻度と汚染レベルの間の相関関係が見つかった[ 16] [ 23]
マサチューセッツ大学アマースト校の動物学教授であったセオドア・デビッド・サージェントは自身の研究(ただし、オオシモフリエダシャクではない 蛾)に基づいてケトルウェルの研究成果に異議を唱えた[ 24]
ケンブリッジ大学遺伝学科の遺伝学者マイケル・マジェラス (英語版 ) [ 25] [ 4] [ 16] [ 4] [ 26] ジェリー・コイン (英語版 ) [ 6] [ 26] [ 27] [ 28] [ 29]
なお、実際にはコインは、オオシモフリエダシャクの工業暗化が自然選択による進化であることは疑っておらず、その自然選択の正体が鳥による捕食だと結論するには不足だと考えていただけであるが[ 30] 科学的な不正 を行ったという根拠のない告発につながっていくことになる。
マイケル・マジェラス]は、ケトルウェルの実験手法に対するさまざまな科学的に正当な批判を解決するために、より精巧な実験を2000年に設計し、2001年からから2007年にかけて実施した。
このマジェラスの新たな実験は、ケトルウェルの実験に関して「蛾の密度が高くなりすぎたのではないか」「昼間に放たれた蛾はうまく隠れ場所を見つけられなかったのではないか」「捕獲された野生の蛾と、研究室育ちの蛾で行動が異なるのではないか」「他の地域から連れてこられた蛾は、本来と異なる行動をとるのではないか」といった疑問に答えるため、より厳密にデザインされた。
2001年から2007年にかけてケンブリッジで行われた研究において、マジェラスは蛾が自然に止まる位置を記録した。実験に使われた135匹の蛾の半分以上が枝に止まり、そのほとんどは枝の下部で、35%が木の幹でほとんどが北側に、13%が小枝の上下に止まった。
ジュディス・フーパーとのやりとりの後、マジェラスはコウモリによる捕食が結果に影響を与えたかどうかを調べる実験を追加した。コウモリは視覚ではなくエコーロケーションによってエサを追いかける。そしてコウモリはどちらの色の蛾も等しく捕食することがわかった。マジェラスはまた蛾を食べている鳥の数種を観察し、これらのデータ全体から、彼は鳥の差別的な捕食がケンブリッジの調査期間における暗化個体の相対的な頻度低下の主要因であると結論した[ 16] [ 31]
実験は最終的に2007年にかけて実施された。マジェラスの死後、この研究結果はイギリスとアメリカの遺伝学者によって解析され、2012年に論文として発表された[ 5] [ 5] ジェリー・コイン (英語版 ) [ 9]
オオシモフリエダシャクのストーリーは小進化を示しており、種分化や大進化を示しているわけではない[ 32] [ 26] [ 33] [ 34] [ 35]
進化の事例としてのオオシモフリエダシャクは反進化論者(創造論者 やインテリジェント・デザイン 支持者など)が好んで取り上げ、進化論を否定しようとする題材の一つである。
反進化論者がオオシモフリエダシャクを気に掛ける理由として、マイケル・マジェラスは「オオシモフリエダシャクのストーリーは理解しやすい。それが反進化論者がオオシモフリエダシャクのストーリーをロビー 攻撃する理由だ。彼らは、多くの人が理解できてしまうだろうということを恐れている」と考察している[ 16]
米国の機関「National Center for Science Education 」の元広報プロジェクトディレクターであるニック・マツケ (英語版 ) [ 26]
ジェリー・コインは、ケトルウェルの実験を、鳥による捕食だと結論するには不足だとして批判した[ 6] [ 30]
オオシモフリエダシャクへの批判は、進化論そのものを学校教育から排除する材料として使われることもある(参考:進化論裁判 )。2000年11月28日、アメリカ・カンザス州プラット郡の教育委員会は、ジョナサン・ウェルズ ほかインテリジェント・デザイン支持者による「Of Pandas and People 」(この本の中でオオシモフリエダシャクにおけるケトルウェルの実験は詐欺的で演出されていると書かれている)のような、インテリジェント・デザイン 支持者から推奨される代替教材(進化を否定する)を使うよう要求することを採択した[ 36] [ 37] [ 38] [ 39]
多くの教科書で用いられている、幹に張り付いたオオシモエダシャクの写真は準備されたものである(死んだ蛾を並べて幹にピン留めしてある)。このことは、「研究すべてが演出されている」という考えと結びつけられた。インテリジェントデザインの指導者フィリップ・E・ジョンソン が、「くさび戦略 (英語版 ) [ 40] ジョナサン・ウェルズ が著書進化のイコンの中で[ 7]
しかしこれらの主張は誤解に基づいたものである。個々の要素を調べるための多くの異なる実験のうちの一つでは、死んだ蛾が木に貼り付けられたものもあったが、その実験は「鳥による捕食率の差」仮説が正しいかどうかを確認するためのものではなく、蛾の密度が鳥の採餌行動にどのような影響が与えるかを調べたものである[ 39] [ 26] [ 26]
生物学者ジェリー・コイン (英語版 ) マイケル・マジェラス (英語版 ) [ 41] [ 42] フィリップ・E・ジョンソン は「くさび戦略 (英語版 ) [ 40] ジョナサン・ウェルズ もまた著書の中で繰り返しこの主張を展開した[ 7] [ 8]
これらは単純に事実と異なる。実際にはマジェラスの著書には、マジェラスが自然状態で休息場所を確認したオオシモフリエダシャク47匹のうち、12匹が木の幹の上におり(6匹はむき出しで、6匹は隠れて)、20匹は枝の付け根に、15匹は枝に止まっていたと記されていた[ 4] [ 26] [ 5]
なお、昆虫学者ドナルド・フラックとのやり取りにおいて、ジョナサン・ウェルズは上述の著書の中でマジェラスが6匹がむき出しの木の幹に止まったと記録していることを認めたが「数万匹の蛾の中で6匹は取るに足りない割合だ」と主張した[ 43] [ 43]
サンデーテレグラフ (英語版 ) [ 42]
本の著者でありマシューズの取材も受けたマジェラスは、この認識が間違いであることを示唆し、またマシューズの記事には科学的な誤り、誤引用、不正確な表現が多いことを指摘しつつ、しかしサイエンスに関する報道の多くはそういうものだと述べた[ 28] [ 28]
ザ・サイエンティスト誌[ 44] ジョナサン・ウェルズ は「C.A.クラークと同僚は25年間のフィールドワークで木の幹に止まっているオオシモフリエダシャクを発見したのはたった一匹だけだった」と書き、「オオシモフリエダシャクが通常木の幹に止まらないという事実はケトルウェルの実験を反証する」と結論した[ 8]
ウェルズはこの中で「教科書が説明しないのは、1980年代以来、生物学者はこの古典的な物語には深刻な欠陥があると気づいていたということだ。もっとも深刻なのは自然のオオシモフリエダシャクは木の幹に止まることすらないということだ。教科書の写真は結局のところ、演出されていた」と述べた[ 7]
Of moths and Man 」(2002年)フーパーは、ケトルウェルが仮説に合致するようデータを改竄し科学的不正を行ったと主張した[ 45]
マジェラスの本のレビューにおいてケトルウェルの実験について批判したコインは、この本でもレビューを書き、オオシモフリエダシャクの工業暗化が自然選択による進化であることを強調し、その自然選択の正体が鳥による捕食だと結論するには不足だと考えていただけであるという立場を改めて表明した上で、フーパ―を「薄っぺらな陰謀説を提出している」と評した[ 30] [ 28] [ 46] [ 47]
^ a b c d e 岩波 生物学辞典 第4版
^ a b c d e Miller, Ken (1999). The Peppered Moth: An Update
^ Natural Selection and Mutation: The Case of the Peppered Moth ^ a b c d Majerus, M.E.N. (1998). Melanism: Evolution in Action. . Oxford University Press, New York.. ISBN 978-0198549833
^ a b c d e Cook, L. M.; Grant, B. S.; Saccheri, I. J.; Mallet, James (2012). “Selective bird predation on the peppered moth: the last experiment of Michael Majerus” . Biology Letters 8 (4): 609–612. doi :10.1098/rsbl.2011.1136 . PMC 3391436 . PMID 22319093 . https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3391436/ .
^ a b c
Coyne, Jerry A. (1998-11). “Not black and white” (英語). Nature 396 (6706): 35–36. doi :10.1038/23856 . ISSN 0028-0836 . http://www.nature.com/articles/23856 .
^ a b c d Wells J. (2000). Icons of Evolution : Science or Myth? Why Much of What We Teach About Evolution is WrongIconsofevolution.com )
^ a b c Wells, J. (1999). "Second Thoughts about Peppered Moths; This classical story of evolution by natural selection needs revising ." The Scientist 13 , 11 (p. 13, May 24, 1999)
^ a b Coyne, Jerry (2012年2月10日). “The peppered moth story is solid ” (英語). Why Evolution Is True . 2021年7月28日 閲覧。
^ Saccheri, I. J.; Rousset, F.; Watts, P. C.; Brakefield, P. M.; Cook, L. M. (2008). “Selection and gene flow on a diminishing cline of melanic peppered moths” . Proceedings of the National Academy of Sciences 105 (42): 16212–16217. doi :10.1073/pnas.0803785105 . PMC 2571026 . PMID 18854412 . https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2571026/ . ^ a b Edleston, R. S. (1864). “Amphydasis betularia”. Entomologist 2 : 150.
^ a b Steward, R.C. (1977). “Industrial and non-industrial melanism in the peppered moth Biston betularia (L.)”. Ecological Entomology 2 : 231–243. doi :10.1111/j.1365-2311.1977.tb00886.x .
^ http://www.talkorigins.org/faqs/wells/images/grantfile.jpg ^ a b Cook LM. (2003). “The rise and fall of the Carbonaria form of the peppered moth.journal = Q Rev Biol. = 78”. The Quarterly Review of Biology 78 (4): 399–417. doi :10.1086/378925 . PMID 14737825 .
^ Tutt, J. W. (1896). British moths . George Routledge and Sons ^ a b c d e f Michael E. N. Majerus (2007年8月). “The Peppered Moth: The Proof of Darwinian Evolution ” (PDF). 2011年6月15日時点のオリジナル よりアーカイブ。2007年9月9日 閲覧。
^ Fisher, R.A. (1933). "On the Evidence Against the Chemical Induction of Melanism in Lepidoptera.", Proceedings of the Royal Society of London, B 112 (pp. 407–416).
^ Bowater, W (1914). “Heredity of melanism in Lepidoptera”. Journal of Genetics 3 : 299-315. doi :10.1007/BF02981796 . ^ Grant BS. (2004). “Allelic melanism in American and British peppered moths..journal = J. Hered”. Journal of Heredity 95 (2): 97–102. doi :10.1093/jhered/esh022 . PMID 15073224 . ^ van't Hof, Arjen E.; Campagne, Pascal; Rigden, Daniel J.; Yung, Carl J.; Lingley, Jessica; Quail, Michael A.; Hall, Neil; Darby, Alistair; Saccheri, Ilik J. (2016). “The industrial melanism mutation in British peppered moths is a transposable element”. Nature 534 (7605): 102–117. doi :10.1038/nature17951 . ^ a b Dobzhansky, T.G. (1937). Genetics and the Evolutionary Process . Columbia University Press. ISBN 0-231-08306-8
^ Haldane, J.B.S. (1924). “A Mathematical Theory of Natural and Artificial Selection”. Transactions of the Cambridge Philosophical Society 23 : 19-41. ^ Young, M. (2003). Moonshine: Why the Peppered Moth Remains an Icon of Evolution .
^ Sargent, T.D., Millar, C.D., and Lambert, D.M. (1998). "The 'classical' explanation of industrial melanism: Assessing the evidence.", Hecht and Wallace (Eds.), Evolutionary Biology volume 23, chapter 9.
^ 英語: Melanism: Evolution in Action.
^ a b c d e f g Nick Matzke (2002-2004). “Icon of Obfuscation ”. Jonathan Wells's book Icons of Evolution and why most of what it teaches about evolution is wrong . TalkOrigins Archive . 2021年8月7日 閲覧。
^ Donald Frack (30 March 1999). “Evolution - March 1999: Peppered Moths - in black and white (part 2 of 2) ”. 2007年9月30日時点のオリジナル よりアーカイブ。2021年8月5日 閲覧。 ^ a b c d Michael Majerus (2004年). “The Peppered moth: decline of a Darwinian disciple ” (.doc). 2007年9月26日時点のオリジナル よりアーカイブ。2021年8月7日 閲覧。
^ Donald Frack (30 March 1999). “Evolution - March 1999: RE: Peppered Moths - in black and white (part 1 of 2) ”. 2007年8月26日 閲覧。 ^ a b c Coyne, Jerry A. (2002-07). “Evolution under pressure” (英語). Nature 418 (6893): 19–20. doi :10.1038/418019a . ISSN 0028-0836 . http://www.nature.com/articles/418019a .
^ Steve Connor, Science Editor (2007年8月25日). “Moth study backs classic 'test case' for Darwin's theory - Independent Online Edition > Sci_Tech ”. The Independent. 2008年10月7日時点のオリジナル よりアーカイブ。2007年9月9日 閲覧。 ^ Tommy Mitchell (2008年4月23日). “Much Ado About Moths ”. 2021年7月31日 閲覧。 ^ Mark Isaak: “Claim CB910.2:Peppered moths are commonly claimed as evidence of evolution, but falsely so. The moths only changed color; they remain peppered moths. ”. The TalkOrigins Archive (2004年). 2021年8月2日 閲覧。 ^ Futuyma, Douglas J.『Evolutionary biology 』(3rd ed)Sinauer Associates、Sunderland, Mass.、1998年、477-478頁。ISBN 0-87893-189-9 。OCLC 37560100 。https://www.worldcat.org/oclc/37560100 。 ^ “Macroevolution: Its definition, Philosophy and History ”. 2008年5月22日 閲覧。 ^ Molleen Matsumura. “Intelligent Design in Pratt County, Kansas ”. National Center Science Education . 2007年8月28日 閲覧。 ^ 英語: Pratt Tribune
^ Coyne, Jerry (2000年12月6日). “Criticism of moth study no challenge to evolution ”. 2021年8月7日 閲覧。 ^ a b Bruce Grant (December 13, 2000). “Charges of Fraud Misleading ”. Pratt Tribune (Kansas). 2008年2月18日 閲覧。
^ a b Donald Frack (16 April 1999). “Evolution - April 1999: Peppered Moths and Creationists ”. 2007年8月26日時点のオリジナル よりアーカイブ。2021年8月7日 閲覧。
^ 英語: Melanism: Evolution in Action.
^ a b Matthews, R. (1999年3月14日). “Scientists pick holes in Darwin moth theory”. The Sunday Telegraph
^ a b Donald Frack (16 April 1999). “Evolution - April 1999: RE: My last word ”. 2007年9月27日時点のオリジナル よりアーカイブ。2021年8月7日 閲覧。
^ 英語: The Scientist
^ Hooper, Judith (2002). Of moths and men : an evolutionary tale : the untold story of science and the peppered moth ISBN 0-393-05121-8 . OCLC 50022818 . https://www.worldcat.org/oclc/50022818 ^ Matt Young. “Moonshine: Why the Peppered Moth remains an Icon of Evolution ”. Department of Physics, Colorado School of Mines. 2007年8月28日 閲覧。 ^ Rudge, David Wÿss (2005-07). “Did Kettlewell commit fraud? Re-examining the evidence” (英語). Public Understanding of Science 14 (3): 249–268. doi :10.1177/0963662505052890 . ISSN 0963-6625 . http://journals.sagepub.com/doi/10.1177/0963662505052890 .
ウィキメディア・コモンズには、
工業暗化 に関連するメディアがあります。




{kind=link}